Khalil et al., 2026 de
Research article
Pathogenic Potential of Hemolytic and Non-Hemolytic Plant Growth
Promoting Bacteria in Galleria mellonella tests
Khalil Md. Masudur Rahman 1, 2*, Figueroa-López Alejandro Miguel 3*, Cantú-Soto Ernesto Uriel 4, Zamudio-Aguilasocho Gloria Margarita 1,5, Cordero-Ramírez Jesús Damián 5, Félix-Gastélum Rubén 3, Maldonado-Mendoza Ignacio Eduardo 1
1Departamento de Biotecnología Agrícola, Instituto Politécnico Nacional, Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional (CIIDIR)- Unidad Sinaloa. CP 81101, Guasave, Sinaloa, México.
2Department of Microbiology, School of Biomedical Sciences, Khwaja Yunus Ali University, Sirajganj, Bangladesh.
3Departamento de Ciencias Naturales y Exactas, Universidad Autónoma de Occidente, Unidad Regional Los Mochis. CP 81217, Los Mochis, Sinaloa, México.
4Departamento de Biotecnología y Ciencias Alimentarias, Instituto Tecnológico de Sonora, 5 de febrero 818 sur, Col. Centro, CP 85000 Ciudad Obregón, Sonora, México.
5Universidad Autónoma de Occidente. Av. Universidad s/n, Frac. Villa Universidad, CP.81044, Guasave, Sinaloa, México.
* Ambos autores hicieron contribuciones iguales al desarrollo de este manuscrito.
Correspondencia: imaldona@ipn.mx
Área Temática: Ciencias de las Plantas Recibido: 03 septiembre 2025 Aceptado: 13 enero 2026 Publicado: 23 enero 2025
Cita: Khalil MMR, Figueroa-López AM, Cantú-Soto E, Zamudio-Aguilasocho GM, Cordero-Ramírez JD, Félix-Gastélum F, and Maldonado-Mendoza IE. 2026 Pathogenic Potential of Hemolytic and Non-Hemolytic Plant Growth Promoting Bacteria in Galleria mellonella tests. Bioc Scientia 2(1). https://doi.org/10.63622/RBS.2518 Copyright: © 2024 by the authors. Submitted for possible open access publication under the terms and conditions of the Creative Commons Attribution (CC BY-NC) license (https://creativecommons.org/licenses/by/4.0/). |
Resumen: La hemólisis bacteriana indica virulencia en humanos con lisis parcial o total de glóbulos rojos[a]. En este estudio, se usaron larvas de Galleria mellonella para evaluar la virulencia humana de 31 bacterias antagonistas de patógenos fúngicos, que previamente mostraron hemólisis alfa y gamma en pruebas in vitro en agar sangre. Se eligió Galleria mellonella para evaluar la virulencia debido a ventajas técnicas como bajo coste, disponibilidad, resultados en 2-3 días, ausencia de cuestiones éticas y similitudes funcionales con el sistema inmunitario de mamíferos e insectos. Se planteó la hipótesis de que las bacterias antagonistas que muestran actividad de lisis parcial (α) y sin lisis (ɣ) en la prueba de agar sangre ejercerán efectos de no virulencia en la prueba in vivo con Galleria. De las 31 cepas bacterianas, tres bacterias productoras de hemolisina γ (Rhizobium pakistanense PaMR4, Rhizobium pakistanense RpCR1 y Staphylococcus saccharolyticus SC4) mostraron virulencia, y las otras veintiocho (hemolíticas α y γ) mostraron efectos no virulentos. La prueba de Galleria sugiere que las bacterias productoras de hemolisina ɣ pueden ser patógenas para los seres humanos, incluso si no pueden lisar los glóbulos rojos (RBC). Por lo tanto, es esencial realizar pruebas de virulencia adicionales a las bacterias que previamente mostraron hemólisis α y ɣ. En este estudio, las 31 cepas bacterianas utilizadas mostraron diferentes respuestas a antibióticos. Algunas cepas Gram-positivas mostraron resistencia a 23 antibióticos, y las cepas Gram-negativas mostraron resistencia a nueve antibióticos. Las cepas bacterianas antagonistas con hemólisis γ, pese a no mostrar actividad hemolítica, podrían mantener capacidad patogénica en humanos. Por lo tanto, se recomienda aplicar pruebas complementarias de virulencia, como el modelo in vivo con Galleria mellonella, para evaluar su bioseguridad. Si se confirma virulencia, deben excluirse como agentes de control biológico en manejo fitosanitario.
Palabras clave: Bacteria antagonista, Galleria mellonella, Hemólisis, Patógeno, Virulencia.
Abstract: Bacterial hemolysis indicates virulence in humans with partial or total lysis of red blood cells. In this study, Galleria mellonella larvae were used to evaluate the human virulence of 31 fungal pathogen antagonist bacteria, which previously showed alpha and gamma hemolysis in in vitro blood agar tests. Galleria mellonella was chosen to evaluate virulence due to technical advantages such as low cost, availability, results in 2-3 days, absence of ethical issues, and functional similarities with the mammalian and insect immune systems. It was hypothesized that antagonistic bacteria showing partial lysis (α) and no lysis (γ) activity in the blood agar test would exert non-virulence effects in the in vivo test with Galleria. From the 31 bacterial strains, three gamma (γ) hemolysin-producing bacteria (Pseudomonas aeruginosa PaMR4, Rhizobium pakistanense RpCR1, and Staphylococcus saccharolyticus SC4) showed virulence, and the other twenty-eight (α and γ hemolytic) showed non-virulent effects. The Galleria test suggests that γ hemolysin-producing bacteria may be pathogenic to humans, even if they cannot lyse red blood cells (RBCs). Therefore, it is essential to perform additional virulence tests on bacteria that previously showed α and γ hemolysis. In this study, the 31 bacterial strains used showed different responses to antibiotics. Some Gram-positive strains showed resistance to 23 antibiotics, and Gram-negative strains showed resistance to nine antibiotics. Antagonistic bacterial strains with gamma hemolysis (γ), despite not showing hemolytic activity, could retain pathogenic capacity in humans. Therefore, complementary virulence tests, such as the in vivo model with Galleria mellonella, are recommended to assess their biosafety. If virulence is confirmed, they should be excluded as biological control agents in phytosanitary management.
Keywords: Antagonistic bacteria, Galleria mellonella, Hemolysis, Pathogen, Virulence.
INTRODUCCIÓN
Chemical fertilizers can contaminate soil, water, and food supply, but biological fertilizers offer a sustainable alternative by promoting plant growth, increasing yield, and improving soil health (Rahimi et al., 2019). Biological fertilizers contain bacteria that promote plant growth by secreting metabolites, hormones, solubilizing minerals, fixing nitrogen, and protecting plants from pathogens (Poria et al., 2022). Plant growth-promoting bacteria in food crops could threaten human health because of virulence factors in their genomes, which can potentially cause diseases. Some bacterial species that can cause human diseases include Acinetobacter baumannii, Bacillus spp, Campylobacter jejuni, Pseudomonas aeruginosa, and Vibrio parahaemolyticus (Fedhila et al., 2006; Peleg et al., 2009; Peleg et al., 2009; Senior et al., 2011; Beeton et al., 2015; Wagley et al., 2018).
In this work, several microorganisms with antagonistic activity against plant fungal pathogens were studied regarding their potential pathogenicity to humans. They belong to the genera Acinetobacter, Bacillus, Pseudomonas, Rhizobium, and Staphylococcus. The Acinetobacter genus comprises the Acinetobacter baumannii complex, containing A. baumannii, A. nosocomialis, A. pitii, and A. calcoaceticus, the most relevant species due to their clinical implications (Nurjadi & Boutin, 2022). It is widely acknowledged that multidrug-resistant Acinetobacter baumannii strains are responsible for infections associated with elevated morbidity and mortality rates (Sunenshine et al., 2007). Bacillus thuringiensis is part of the Bacillus cereus group (Bacillus cereus sensu lato). This group includes 21 species, some of which have been reported for their importance in the medical field (Baek et al., 2019). Pseudomonas aeruginosa is another human pathogen implicated in pulmonary infections (Vidaillac & Chotirmall, 2021), as well as the Staphylococcus genus, where there are some species reported with clinical importance, including the most common S. aureus and S. saccharolyticus that can cause human illness (Wang et al., 2020).
Virulence factors in bacteria result from genes expressed via transcription and translation, often located in chromosomes but sometimes in plasmids. These factors enable bacteria to complete their infective cycle in hosts, including humans, animals, or plants. Typically, these proteins or enzymes, called virulence factors, are key in host cell invasion, evading defenses, and toxin production that cause disease (Leitão, 2020).
Certain bacteria within the same genus and species show different pathogenic behaviors due to their virulence factors. Even within the same species, some vary in the virulence factors needed to cause disease (Leitão, 2020). Virulence factors like adhesion help bacteria complete their infective cycle factors. Some pathogenic bacteria produce fimbriae for adherence, invasion factors that facilitate cell entry, and are surrounded by capsules that protect from opsonization and phagocytosis. Additionally, toxin production is crucial since Gram-negative bacteria's lipopolysaccharide endotoxins cause fever, blood pressure changes, inflammation, and shock. Endotoxins are protein toxins or enzymes secreted by pathogens, including cytotoxins, neurotoxins, and enterotoxins (Leitão, 2020).
If all virulence factors are produced, a successful infection by the pathogen can develop. It is known that the malfunction of any of them can lead to a reduction in pathogenicity or completely inhibit it. In Listeria monocytogenes EGD-e, a mutation in the hlyA gene (the gene responsible for the production of listeriolysin O) renders the bacteria completely avirulent, resulting in the complete absence of infection in chicken embryos (Quereda et al., 2018). There is a naturally occurring strain of Listeria monocytogenes classified as ATCC 15313, which is avirulent because it cannot produce listeriolysin O. These are examples of bacteria of the same species that can exhibit variations in their pathogenic behavior, depending on their virulence factors. Therefore, this paper hypothesizes that hemolysis analysis on blood agar is not decisive in ruling out or approving the use of microorganisms in crops of interest for food production. To prevent this, microorganisms used as biotechnological products against plant diseases and fungal infections in agriculture must be thoroughly evaluated and tested to mitigate risks to human health. A viable alternative is the use of eukaryotic models such as Galleria mellonella. Pathogenicity analysis in eukaryotic models is essential to provide the necessary evidence of the pathogenic behavior of microorganisms.
Insects are a successful group of invertebrate animals that serve as a model for studying the virulence of microbial pathogens in humans (Tsai et al., 2016). Though insects (invertebrates) and vertebrates are different animal groups, vertebrates have developed an adaptive immune response. In contrast, the vertebrate innate immune system retains strong structural and functional similarities to the insect immune system (Browne et al., 2013). A group of insects, including Galleria mellonella (greater wax moth), Drosophila melanogaster (fruit fly), Manduca sexta (tobacco hornworm or Goliath worm), and Bombyx mori (silkworm), as well as, Danio rerio (zebra fish) are widely considered alternative model organisms, and the virulence results obtained in these model organisms are similar to those obtained with mammals (Fuchs & Mylonakis, 2006; Browne et al., 2013).
The genus Galleria is part of the Pyralidae family. The larvae of Galleria mellonella are used as a model system for testing bacterial virulence in vivo. A few features considered for G. mellonella larvae as a model test include availability, low cost, basic requirements, and the possibility of obtaining results in a short time period, such as two to three days (Desbois & Coote, 2012; Kay et al., 2019). This insect's larvae can be rapidly grown at 30–37° C, enabling scientists to examine the temperature-dependent microbial virulence factors (Fallon et al., 2012). In addition to these technical advantages, G. mellonella larvae are the preferred choice for testing bacterial pathogenicity in humans (Kavanagh y Fallon, 2010; Salgado-Morales et al., 2019). Recent studies have shown that agricultural ecosystems are key reservoirs for potential human infections, highlighting the need for vigilant surveillance to anticipate disease emergence. Some plant pathogens are opportunistic and can infect humans through cross-infections. Focus on understanding pathogenicity mechanisms to reduce cross-infection risks (Kim et al., 2020).
For these reasons, it is also hypothesized that antagonistic bacterial strains that exhibit α (with partial lysis effects) and γ-hemolysin (without lysis effects) are expected to be non-virulent, although there may be exceptions, and these will not be pathogenic in the eukaryotic model Galleria mellonella. In alignment with our hypothesis, we investigated the virulence of several plant growth-promoting bacteria to discard potential human virulent bacteria. Furthermore, this study examined the antibiotic resistance of antagonistic bacterial strains.
MATERIALS AND METHODS
Preparation of biological materials
Thirty-one antagonistic bacterial strains were evaluated for their virulence effects on humans by in vivo Galleria mellonella tests (Table 1). Six bacterial strains Escherichia coli O157-H7 28-1 (Rivas-Ruiz et al., 2020), Escherichia coli O157-H7 (Rivas-Ruiz et al., 2020), Escherichia coli JA 31 EC (Anduro-Jordan et al., 2022) Listeria monocytogenes ATCC 7644, Staphylococcus aureus ATCC11532 and Salmonella sp ATCC 14028 were used as positive controls and a non-pathogenic Escherichia coli ATCC 25922 was used as a negative control. The Microbiology Laboratory of the Department of Biotechnology and Food Science at Instituto Tecnológico de Sonora (ITSON), Sonora, Mexico, provided the control bacteria. Five of the 31 bacterial strains (AcDB3, BaMA26, BsTA16, BsiDA2, and BtMB9) were obtained from three bacterial collections deposited at the Department of Agricultural Biotechnology at CIIDIR-Sinaloa, Instituto Politécnico Nacional (Mexico) previously described in Khalil et al., (2021). The other 26 bacterial strains (ArCR5, BaCR3, BvCR6, BvML3, CL1, CL2, CL3, CL4, CL5, CL6, CL7, CL8, CS1, CS2, CS3, CS4, ML2, MR1, MS1, PaMR4, PgML1, PpMR2, PpMR3, RpCR1, SsCR4 and SwCR2) were isolated from the rhizosphere of giant reed (Arundo donax L.) and maize (Zea mays L.). The use of plant growth-promoting microorganisms is crucial for enhancing agricultural sustainability. These organisms, including those yet to be molecularly characterized in this work, represent promising candidates and have demonstrated benefits to plants by augmenting nutrient availability and providing protection against fungal pathogens (Khalil et al. 2021).
All bacterial strains were maintained in Luria Bertani broth (LB, Sigma, No. Cat. L3022, USA) supplemented with 15% glycerol at -80 °C. To check the viability of the strains, all cryopreserved bacteria were transferred onto LB agar (LBA) and incubated at 30 °C for 24 h. Nine bacterial strains (BaMA26, BsTA16, BsiDA2, CL1, CL4, CL7, CL8, CS3 and CS4) required the addition of glucose (1% w/v) with LBA for reactivation and growth. After growing bacteria in LBA, a single colony was transferred to 5 mL of LB broth and incubated at 30 °C for 24 h at 200 rpm. Next, 1 mL of each bacterial suspension (AcDB3, BaMA26, BsTA16, BsiDA2, and BtMB9) was transferred to 100 mL of LB and incubated at 30 °C for 9 h at 200 rpm (Cordero-Ramírez et al., 2013).
Hemolysis tests on blood agar medium
Bacteria were grown overnight in 5 mL of LB medium at 30 °C at 200 rpm. Briefly, 1 mL of bacterial suspension was transferred to a 1.6 mL microcentrifuge tube and centrifuged at room temperature for 20 min at 13,000 rpm. Next, 50 µL of supernatant was added to 5 mm circular wells on the blood agar medium. The wells were previously made using a sterile rubber plug punch in the blood agar medium at 5% of sheep’s blood (MCD Lab, Cat. 7504). Plates were then incubated for 24 h at 37 °C. The results were interpreted based on the clear zone surrounding the wells: β-hemolysis was observed as a clear zone, demonstrating complete breakage of erythrocytes; α-hemolysis was revealed by a slight change in color surrounding the wells, indicating partial erythrocytes breakdown; and γ-hemolysis (or no hemolysis) was suggested by the absence of any change in color or clearness of the medium surrounding the well (Misawa et al., 1995). Bacteria showing β-hemolysis were discarded, and only α and γ-hemolytic bacteria were used in subsequent studies.
Virulence assays in Galleria mellonella
Galleria mellonella (G. mellonella) larvae were used as a model to detect the virulence of 31 potential antagonistic bacterial strains (Table 1) against humans, as described by Menard et al. (2021). The G. mellonella eggs were grown in a plastic container and fed. The larvae were fed pellets that had been previously prepared. Briefly, 50 g of honey with 50 mL of H2O, 90 g of maize powder, 50 g of wheat powder, 50 g of milk powder, 20 g of yeast, 1.2 g of sodium benzoate, 50 mL of glycerol, and 50 mL of distilled water were placed in a beaker. Next, the mixture was boiled for 5 min at 100 °C. All ingredients were mixed, made into a mold, and put into a small plastic container with insect larvae. Larvae were maintained at 30 °C for 60 days. A plastic sealing film was applied to the top of the container, and small perforations were made in the plastic seal to facilitate oxygen exchange. The holes were secured with micropore tapes to prevent the small Galleria larvae from passing through the pores. When the caterpillar has grown sufficiently to be handled, it is transferred to a new container and classified by size. Only the caterpillars in the last larval stage were used for the experiments (Ménard et al., 2021).
The bacterial strains were incubated overnight at 30 °C in LBA and or LBGA medium, depending on bacterial growth conditions. The overnight cultured bacteria were inoculated in 5 mL of LB broth medium and incubated for 12 h at 30 °C. After 12 h of incubation, 50 µL of bacterial suspension was placed in 5 mL of LB broth medium and incubated at 30 °C for 6 h. Subsequently, the bacterial pellets were collected by centrifugation at 10,000 rpm for 1 min. One mL of MgSO4•7H2O was added into a tube containing bacterial pellets and made a serial dilution up to 10-6 using MgSO4•7H2O (Faga Lab, Cat. 2344). Two bacterial dilutions, 10-5 and 10-6, were used for injecting into Galleria larvae and the colony forming units (CFU) were counted. Twenty µL of pellet suspension from each dilution [(10-5 (500 CFU) and 10-6 (50 CFU)] were injected into each Galleria larvae using a 0.3 mL U-100 (DL ®) insulin syringe, and 50 µL of them were spread on LBA and or, LBGA medium and incubated at 30 °C overnight and the bacterial CFU were counted. Infected Galleria larvae were maintained at 30 °C for 48 hours, and mortality was recorded at 24 hours. The assay was performed three times, using ten Galleria larvae for each bacterial test.
Antimicrobial susceptibility test
An analysis of the minimum inhibitory concentration (MIC, μg/mL) was performed on MicroScan autoSCAN-4 System Beckman Coulter® to determine susceptibility or resistance to antibiotics, according to the Clinical and Laboratory Standards Institute (CLSI) guidelines for the antibiotics listed in Tables 2 and 3, for Gram-negative and Gram-positive bacteria, respectively (CLSI, 2024).
RESULTS AND DISCUSSION
Bacterial hemolysis on blood agar tests and mortality effects on Galleria mellonella model
The present study examined the virulence and antibiotic resistance of 31 plant rhizospheric bacteria on eukaryotic models. These bacteria show potential for combating plant pathogens and promoting plant growth. Alpha (α)-hemolysin and γ -hemolysin bacteria were chosen as antagonists, showing minimal or no blood agar color change, indicating low erythrocyte lysis. Eighteen (AcDB3, BaMA26, BtMB9, CL1, CL2, CL3, CL4, CL5, CL6, CL7, CL8, CS1, CS2, CS3, CS4, ML2, MR1 and MS1) out of 31 bacteria showed partial hemolysis (α- lysis) and 13 strains (ArCR5, BaCR3, BsiDA2, BsTA16, BvCR6, BvML3, PaMR4, PgML1, PpMR2, PpMR3, RpCR1, SsCR4 and SwCR2) showed γ hemolysis (no hemolysis) (Table 1).
Table 1. Galleria virulence test performed on bacteria isolated from different plant rhizospheres with antagonistic and plant growth-promoting effects.
NI = non-identified. Gamma (γ)- hemolysis = no erythrocyte lysis, alpha (α)- hemolysis = slight erythrocyte lysis, beta (β)- hemolysis = complete erythrocyte lysis. (*) Control strains.
Based on the hypothesis that hemolytic microorganisms are a health risk in food production (Gera y McIver, 2013), the hemolysis test excludes microorganisms with partial or complete hemolysis on blood agar plates (Patel, 2023). The results suggest that blood lysis is not a key factor in outcomes since human infection needs specific virulence factors.
Acinetobacter calcoaceticus, part of the A. calcoaceticus-baumannii complex, is an opportunistic bacterium causing pneumonia and bacteremia (Mancilla-Rojano et al., 2020). Despite this, A. calcoaceticus isolates from this work is not a bacterium of significant medical importance, as it is only slightly associated with human infections. Nevertheless, the presence of phospholipases within its genome has the potential to catalyze the breakdown of phospholipids in red blood cell membranes, which can lead to partial hemolysis (Table 1) (Lehmann, 1973).
It is reported that isolates belonging to the species of Bacillus amyloliquefaciens do not adversely affect human health and have been used as a probiotic against Clostridium perfringens (Chen et al., 2024). However, in this work, B. amyloliquefaciens showed alpha hemolysis, and it is reported that this bacterium can produce phospholipases, which are responsible for the degradation of phospholipids, thereby explaining the alpha hemolysis behavior (Chen et al., 2024).
Bacillus thuringiensis is a well-studied bacterium with a wide range of applications in agriculture. It also produces phospholipases responsible for lysing erythrocytes, resulting in partial blood hemolysis in isolates of this study (Lereclus et al., 1996).
The results obtained for Pseudomonas aeruginosa suggest that this strain cannot exert hemolysis on the blood agar assay, possibly due to the lack of the gene that encodes phospholipase C (Wolfmeier et al., 2022). However, P. aeruginosa retains its pathogenicity in the absence of phospholipases, but its potential to induce disease may be reduced. Although phospholipases represent certain virulence factors that facilitate tissue damage and advance disease progression, they do not constitute the only elements that contribute to the bacterium's pathogenicity. Other virulence factors, including exotoxins, proteases, and adhesins, also play a significant role in the infections caused by P. aeruginosa (Strateva & Mitov, 2011).
The Rhizobium genus is a group of bacteria closely associated with the colonization and nodulation of plants. There are few reports of human infection by Rhizobium species. The species reported to be pathogenic to humans is Rhizobium pusense (Aujoulat et al., 2015). However, there are no documented cases of Rhizobium pakistanense infecting humans. In the present study, R. pakistanense RpCR1 did not exhibit hemolytic activity in the blood agar tests. This phenotypic characteristic may be associated with the absence of hemolytic or proteolytic enzymes that can affect the erythrocyte cell membrane.
Staphylococcus saccharolyticus is a recently described human pathogen that acts as a skin colonizer (Ahle et al., 2020), but its pathogenicity mechanism is unclear (Brüggemann et al., 2019). The results suggest that Staphylococcus saccharolyticus SsCR4 was unable to degrade erythrocytes on blood agar, suggesting that it could not produce proteins capable of lysing red blood cells.
Galleria mellonella virulence assay shows that non hemolytic bacterial strains may show the ability to behave as virulent.
It was observed that non-hemolytic biological control agents showed virulence in the in vivo G. mellonella test (Table 1). It also supports previous studies indicating that G. mellonella is an easy, cost-effective, and successful model system for evaluating bacterial virulence against humans. The Galleria test also demonstrated that all α-hemolytic bacteria tested were non-pathogenic, but some γ-hemolytic bacteria tested showed virulence, which partially contradicts our hypothesis that the α- and γ-hemolytic bacteria will behave as non-virulent in this test (Table 1).
The insect G. mellonella has been used to inspect bacterial virulence against human infectious agents such as diarrheal E. coli (Khalil et al., 2016) and against the fish pathogen Vibrio anguillarum (causes inflammation and skin disease) (McMillan et al., 2015). G. mellonella has also been used to test bacterial virulence against humans, but so far, it has not been reported that γ-hemolytic plant rhizospheric bacteria can be pathogenic to humans.
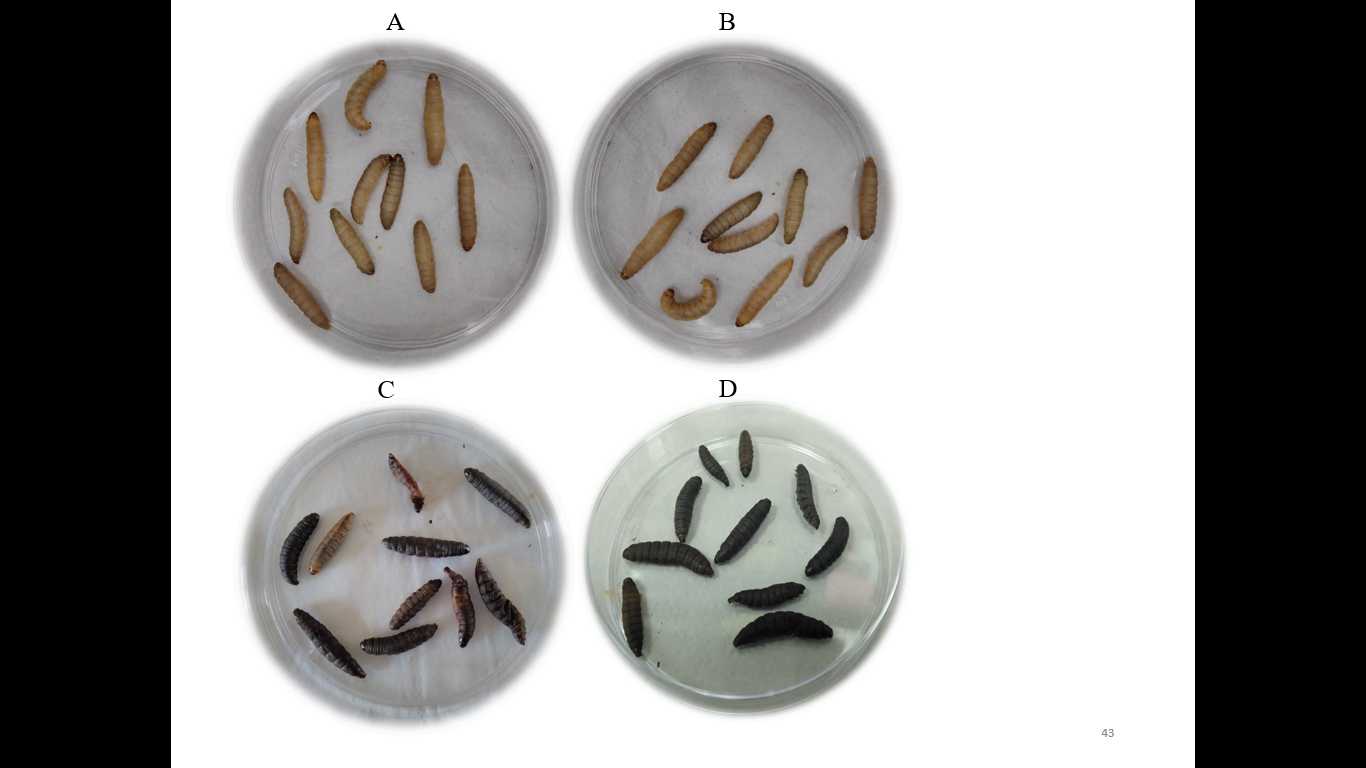
The six human pathogenic strains injected into Galleria as positive controls showed 100% mortality in both concentrations (50 and 500 CFU per Galleria) (Table 1 and Figure 1 D). After 24 h of injection, the Galleria mortality was 30 to 40% using 50 CFU and 50 to 80% with 500 CFU (Table 1). The non-pathogenic Escherichia coli ATCC 25922 strain injected into Galleria as a negative control did not show mortality at both concentrations after 24 h and 48 h (Table 1 and Figure 1A).
Figure 1. Virulence test of thirty-one antagonistic bacteria was performed in Galleria mellonella insects. The percentage of Galleria mortality was counted after 24 and 48 h of infection period. A non-pathogenic strain of Escherichia coli ATCC 25922 was used as negative, and eight human pathogenic bacteria were used as positive controls. A) The infected Galleria with Escherichia coli ATCC 25922 (negative control) and B) Acinetobacter calcoaceticus AcDB3 (antagonist) were alive after 48 h of incubation. C) The Galleria had died after 48 h of incubation while infected with the antagonistic bacterium of Rhizobium pakistanense RpCR1 (ɣ-hemolysis) and D) Staphylococcus aureus ATCC 11632 (positive control). Ten replicates were used for each bacterium.
Among all the rhizospheric bacteria evaluated, 18 showed α-hemolysis (AcDB3, BaMA26, BtMB9, CL1, CL2, CL3, CL4, CL5, CL6, CL7, CL8, CS1, CS2, CS3, CS4, ML2, MR1, and MS) (Table 1). Bacteria originating from different plant rhizospheres, such as Acinetobacter sp., Bacillus sp., and Pseudomonas aeruginosa, show plant-growth-promotion and antagonism against different phytopathogens (Méndez et al., 2021; Schreiter et al., 2018; Wang et al., 2021). Acinetobacter sp. (Jang et al., 2016) and Bacillus spp. (Escamilla-Montes et al., 2015) are well known as bio-control agents and widely distributed in plant rhizospheres. Biofilm formation is a necessary trait to attach to the plant root surfaces to persist for a long time and work against phytopathogens (Chen et al., 2013). A few species of Acinetobacter (Giannouli et al., 2013) and Bacillus, (Granum, 1994) are the causative agents of human foodborne diseases. Ehling-Schulz et al. (2004) reported that Bacillus cereus carried food toxins that may cause human diseases. Guinebretière et al. (2002) mentioned that B. cereus could secrete four different types of enterotoxins which may cause food-born diarrhea: two protein complexes, hemolysin BL (HBL) and nonhemolytic enterotoxin (NHE), and two enterotoxin proteins: enterotoxin T (bc-D-ENT) (Agata et al., 1995) and cytotoxin K (Lund et al., 2000). Based on the hypothesis that hemolytic bacteria pose a risk to human health, this study indicated that all the alpha-hemolytic bacteria tested did not present any risk in the Galleria mellonella model and exhibited no virulence.
Among all the rhizospheric bacteria evaluated 13 were γ-hemolytic (ArCR5, BaCR3, BsTA16, BsiDA2, BvCR6, BvML3, PaMR4, PgML1, PpMR2, PpMR3, RpCR1, SsCR4 and SwCR2). However, Pseudomonas aeruginosa (PaMR4), Rhizobium pakistanense (RpCR1), and Staphylococcus saccharolyticus (SsCR4) were virulent in the Galleria test (Table 1, Figures B and C). PaMR4 showed 100% mortality in both concentrations after 24 h and 48 h of incubation, while RpCR1 and SsCR4 evaluated at 500 CFU showed 100% mortality and 33.4% mortality after 48 h, respectively (Table 1).
The results suggest that the Galleria test indicates that the pathogenicity of antagonistic bacteria originating from different plant rhizospheres does not depend on their red blood cell (RBC) lytic effects. Furthermore, the results obtained from Galleria tests demonstrated that the virulence of pathogenic bacteria varies, possibly due to the presence or absence of virulence factors and their relative doses in each strain (Bokhari et al., 2017).
Six human pathogenic bacteria were injected into G. mellonella insects, revealing that pathogenic bacteria are detrimental to this eukaryotic model, but their virulence capacity depends on the bacterial doses. A dose of 50 CFU showed a lower mortality than 500 CFU after the same incubation period, which confirmed that bacterial virulence depends on a pathogen dose. Bokhari et al. (2017) reported that the virulence in G. mellonella is influenced by the relative injected dose, and our findings support their statements.
Divyakolu et al. (2019) reported that gamma (γ)- hemolysin-producing bacteria (Staphylococcus aureus) can threaten humans, and our results agree with their findings. In the present study, three antagonistic bacteria (PaMR4, RpCR1, and SsCR4) that did not show hemolysis (γ-hemolysis) in vitro showed virulence in Galleria mellonella tests, indicating that these antagonistic bacteria can possibly cause human disease.
Pseudomonas aeruginosa strain was the most virulent antagonist bacterium (Table 1). Morin et al. (2021) reported that this bacterium causes multiple infectious diseases, such as lung infection with chronic diseases, primary ciliary dyskinesia, and ventilator-associated pneumonia, bacteremia and sepsis, urinary tract infection, and diabetic foot ulcers.
Several authors previously mentioned that a few species of the genus Rhizobium are opportunistic human pathogens. Kuchibiro et al. (2018) reported that Rhizobium could cause sepsis in the human body if the patient is previously affected with diabetes, hyperlipidemia, hypertension, hypothyroidism, and osteoporosis. Rhizobium pakistanense (RpCR1) showed virulence in vivo in the G. mellonella test, demonstrating that this bacterium is possibly pathogenic to humans. Khalid et al. (2015) reported that Rhizobium pakistanense is a novel species in the group of the genus Rhizobium, and it has not been reported as pathogenic to humans until now.
Staphylococcus aureus is a common human pathogenic bacterium that causes typical skin infections in humans and animals (Divyakolu et al., 2019; Hanselman et al., 2009; Wertheim et al., 2005). Staphylococcus warneri is a less common pathogen in humans, but in some cases can cause osteoarticular infection (Legius et al., 2012). Staphylococcus saccharolyticus can cause different human diseases, such as spondylodiscitis (Godreuil et al., 2005; Mikhael et al., 2009; Trojani et al., 2020), endocarditis (Westblom et al., 1990), pyomyositis (Young & Bhally, 2017), pneumonia (Wu et al., 2009), empyema (Wang et al., 2020), and bone marrow infection (Liu et al., 2015). This corresponds with the virulence observed for this strain (Table 1).
The 31 antagonistic bacterial strains used in this study have a strong capacity to control different phytopathogens in vitro or in vivo and enhance plant growth (Khalil et al., 2021; Zamudio-Aguilasocho, 2019). However, the virulence results indicate that PaMR4, RpCR1, and SsCR4 cannot be used as biological control agents against phytopathogens because they represent significant risks to humans and animals.
This research could introduce a new concern in the agricultural crop protection sector, where biological control agents are used against phytopathogens. Hemolysin production is not the only key factor in determining bacterial pathogenicity. Pathogenic bacteria can secrete other virulence factors such as coagulase, enterotoxins, toxic shock syndrome toxin 1, exfoliative toxins, and Panton-Valentine leucocidin. In human pathogenic E. coli, approximately ninety proteins acting as virulence factors with different functions are associated to human disease (Pakbin et al., 2021). Listeria monocytogenes showed over eleven virulence factors reported to be involved in mechanisms like survival, virulence, antimicrobial resistance, and persistence in unfavorable environmental conditions (Vera et al., 2013; Matereke & Okoh, 2020). Thus, human pathogenesis is not exclusively due to hemolysin production and may require a combination of virulence factors.
In conclusion, the three non-hemolytic bacteria, PaMR4, RpCR1, and SsCR4, showed virulence in the Galleria mellonella virulence test indicating they can cause human diseases. Therefore, before using biological control agents to protect crops from devastating diseases, it is essential to confirm their virulence effects with a combination of different virulence tests.
Antagonistic bacterial strains demonstrated resistance to various antibiotic compounds.
All the bacterial strains showed different responses to the antibiotics tested. The gram-negative bacteria resistance ranged from 10 to 23 antibiotics; the highest resistance was observed for strain PaMR4 that showed resistance against 23 out of the 24 antibiotics tested, followed by PgML1 and ML2 that were resistant to 20 antibiotics (Table 2). Other strains with high antibiotic resistance were PpMR2 (17), PpMR3 (15), AcDB3 (10) and ArCR5 (10) (Table 2). For gram-positive bacteria, BtMB9 was resistant to 13 out of 21 antibiotics tested (Table 3). the isolates RpCR1, BvCR6, MS1, SsCR4, SwCR2, and BsTA16 showed resistance to 9, 6, 5, 5, 5, and 4 antibiotics, respectively (Table 3). Thirteen strains (BaCR3, BvML3, CL1, CL2, CL3, CL4, CL5, CL7, CL8, CS1, CS2, CS3, CS4) showed resistance to 3 antibiotics. The strain BaMA26 was resistant to two antibiotics, and BsiDA2 to only one (Table 3).
The unregulated use of antibiotics for animal food production has put a lot of pressure on the industry, which is reflected in the rise in antimicrobial resistance (AMR) that has become a public concern. Furthermore, if resistant microorganisms are used and applied in agricultural food production, they should be handled carefully to reduce the risk of illness.
Pseudomonas aeruginosa was the most virulent bacterial strain (Table 1) and it also showed resistance to 23 out of 24 antibiotics tested (Table 2). This species has previously shown resistance to several antibiotic compounds, such as aminoglycosides, β-lactam agents, and fluoroquinolones (Smith et al., 2016). These microorganisms can resist up to seven classes of antibiotics (Meng et al., 2020). P. guariconensis and P. plecoglossicida strains resisted 20 and 17 antibiotic compounds, respectively (Table 2). Therefore, the present study demonstrated that several Pseudomonas spp. strains are multi-drug-resistant.
Table 2. Antibiotic susceptibility of eight gram-negative bacteria against twenty-four antibiotics.
Strains ID | Antibiotics (M.I.C µg/mL) | |||||||||||||||||||||||
AMC | AMK | AMP | CAZ | CAZC | CFZ | CIP | CRO | CTX | CTXC | CXM | ETP | FEP | FOF | GEN | IPM | LVX | MEM | SAM | SXT | TET | TGC | TOB | TZP | |
AcDB3 | R>16/8 | S<16 | R>16 | S<1 | R>2 | R>4 | S<1 | R>32 | R>32 | R>4 | R>16 | R>2 | S<4 | S | S<2 | S<1 | S<2 | S<1 | S<8/4 | R>2/38 | S<4 | S<2 | S<4 | S<16 |
ArCR5 | R>16/8 | S<16 | R>16 | R>16 | S<0.25 | R>4 | R>2 | R>32 | R>32 | S<0.5 | R>16 | R>2 | S<4 | S | S<2 | S<1 | S<2 | S<1 | S<8/4 | R>2/38 | S<4 | S<2 | S<2 | S<16 |
CL6 | S<8/4 | S<16 | S<8 | S<1 | S<0.25 | S<2 | S<1 | S<1 | S<2 | S<0.5 | S<4 | S<0.5 | S<4 | S | S<2 | S<1 | S<2 | S<1 | S<8/4 | S<2/38 | S<4 | S<2 | S<4 | S<16 |
ML2 | R>16/8 | S<16 | R>16 | S<1 | R>2 | R>4 | R>2 | R>32 | R>32 | R>4 | R>16 | R>2 | R>16 | R | R>8 | R>8 | R>4 | S<1 | R>16/8 | R>2/38 | R>8 | R>4 | S<4 | R>64 |
PaMR4 | R>16/8 | R>32 | R>16 | R>16 | R>2 | R>4 | S<1 | R>32 | R>32 | R>4 | R>16 | R>2 | R>16 | R | R>8 | R>8 | R>4 | R>8 | R>16/8 | R>2/38 | R>8 | R>4 | R>8 | R>64 |
PgML1 | R>16/8 | S<16 | R>16 | S<1 | R>2 | R>4 | R>2 | R>32 | R>32 | R>4 | R>16 | R>2 | R>16 | R | R>8 | R>8 | R>4 | S<1 | R>16/8 | R>2/38 | R>8 | R>4 | S<4 | R>64 |
PpMR2 | R>16/8 | S<16 | R>16 | S<1 | R>2 | R>4 | R>2 | R>32 | R>32 | R > 4 | R > 16 | R>2 | S<4 | S | S<2 | R>8 | R>4 | S<1 | R>16/8 | R>2/38 | R>8 | R>4 | S<4 | R>64 |
PpMR3 | R>16/8 | S<16 | R>16 | S<1 | R>2 | R>4 | S<1 | R>32 | R>32 | R > 4 | R>16 | R>2 | S<4 | S | S<2 | R>8 | S<2 | S<1 | R>16/8 | R>2/38 | R>8 | R>4 | S<4 | R>64 |
The letter R refers to resistance, and S indicates sensitivity. AcDB3 = Acinetobacter calcoaciticus, ArCR5 = Acinetobacter radioresistens, Pa = Pseudomonas aeruginosa, Pg = Pseudomonas guariconensis, Pp = Pseudomonas plecoglossicida. Amoxicillin-clavulanate = AMC, Amikacin = AMK, Ampicillin = AMP, Ceftazidime = CAZ, Ceftazidime-clavulanate = CAZC, Cefazolin = CFZ, Ciprofloxacin = CIP, Ceftriaxone = CRO, Cefotaxime = CTX, Cefotaxime-clavulanate = CTXC, Cefuroxime = CXM, Ertapenem = ETP, Cefepime = FEP, Fosfomycin = FOF, Gentamicin = GEN, Imipenem = IPM, Levofloxacin = LVX, Meropenem = MEM, Ampicillin-sulbactam = SAM, Trimetropim-sulfametoxazol = SXT, Tetracycline = TET, Tigecycline = TGC, Tobramycin = TOB, Piperacillin-tazobactam = TZP. M.I.C = Minimum inhibitory concentration.
Although multidrug resistance is rising, this topic is well studied due to the Bacillus thuringiensis insecticidal applications (Gao et al., 2018). This strain (BtMB9) was identified in the present study as one of the most resistant bacteria (resistant to 13 antibiotics) (Table 3). Bacillus thurigiensis is closely related to Bacillus cereus, and both belong to the Bacillus cereus group. Antibiotic resistance has been studied in species of this group due to their multidrug resistance behavior towards aminoglycosides, β-lactams, cephalosporins, and fluoroquinolones (Gao et al., 2018).
The Acinetobacter genus is of public concern; the representative microbial species is A. baumannii, which belongs to the Acinetobacter baumannii complex, which also includes A. calcoaceticus.
These microbial species have shown multidrug resistance (MDR) to several antibiotic compounds, like aminoglycosides, β-lactams, fluoroquinolones, chloramphenicol, erythromycin, and tigecycline, as in this work (Bello-López et al., 2020).
Resistant bacteria can cause human and animal diseases, and their infections are more severe than those caused by non-resistant bacteria. Antibiotic resistance can lead to higher medical costs, prolonged hospital stays, and significantly increased mortality (WHO, 2020).
Table 3. Antibiotic susceptibility of twenty-three gram-positive bacteria against twenty-one antibiotics.
Strain ID | Antibiotics (M.I.C ug/mL) | ||||||||||||||||||||
AMC | AMP | CIP | CLI | CRO | DAP | ERY | FOF | GEN | LVX | LZD | MXF | NIT | OXA | PEN | RIF | SAM | SXT | SYN | TET | VAN | |
BaCR3 | S<4/2 | S<2 | S<1 | S<0.5 | S<8 | S<0.5 | R>4 | S | S<4 | S<1 | S<1 | S<0.5 | R>64 | S<0.25 | S<0.03 | S<1 | S<8/4 | S<0.5/9.5 | S<0.5 | R>8 | S<0.25 |
BaMA26 | S<4/2 | S<2 | S<1 | R>4 | S<8 | S<0.5 | S<0.5 | S | S<4 | S<1 | S<1 | S<0.5 | S<32 | S<0.25 | S<0.03 | S<1 | S<8/4 | S<0.5/9.5 | R>2 | S<4 | S<0.25 |
BsTA16 | S<4/2 | R>8 | S<1 | R>4 | S<8 | S<0.5 | S<0.5 | S | S<4 | S<1 | S<1 | S<0.5 | S<32 | S<0.25 | R>8 | S<1 | S<8/4 | S<0.5/9.5 | R>2 | S<4 | S<0.25 |
BsiDA2 | S<4/2 | S<2 | S<1 | S<0.5 | S<8 | S<0.5 | S<0.5 | S | S<4 | S<1 | S<1 | S<0.5 | S<32 | S<0.25 | S<0.03 | S<1 | S<8/4 | S<0.5/9.5 | R>2 | S<4 | S<0.25 |
BtMB9 | R>4/2 | R>8 | S<1 | R>4 | S<8 | R>4 | R>4 | R | S<4 | S<1 | R>4 | S<0.5 | S<32 | R>2 | R>8 | R>2 | R>16/8 | S<0.5/9.5 | R>2 | S<4 | R>16 |
BvCR6 | S<4/2 | R>8 | S<1 | S<0.5 | S<8 | S<0.5 | R>4 | S | S<4 | S<1 | R>4 | S<0.5 | S<32 | S<0.25 | R>8 | S<1 | S<8/4 | R>2/38 | R>2 | S<4 | S<0.25 |
BvML3 | S<4/2 | S<2 | S<1 | S<0.5 | S<8 | R>4 | S<0.5 | S | S<4 | S<1 | S<1 | S<0.5 | S<32 | S<0.25 | S<0.03 | S<1 | S<8/4 | S<0.5/9.5 | R>2 | S<4 | S<0.25 |
CL1 | S<4/2 | R>8 | S<1 | S<0.5 | S<8 | S<0.5 | S<0.5 | S | S<4 | S<1 | S<1 | S<0.5 | S<32 | S<0.25 | R>8 | S<1 | S<8/4 | S<0.5/9.5 | R>2 | S<4 | S<0.25 |
CL2 | S<4/2 | R>8 | S<1 | S<0.5 | S<8 | S<0.5 | S<0.5 | S | S<4 | S<1 | S<1 | S<0.5 | S<32 | S<0.25 | R>8 | S<1 | S<8/4 | S<0.5/9.5 | R>2 | S<4 | S<0.25 |
CL3 | S<4/2 | R>8 | S<1 | S<0.5 | S<8 | S<0.5 | S<0.5 | S | S<4 | S<1 | S<1 | S<0.5 | S<32 | S<0.25 | R>8 | S<1 | S<8/4 | S<0.5/9.5 | R>2 | S<4 | S<0.25 |
CL4 | S<4/2 | R>8 | S<1 | S<0.5 | S<8 | S<0.5 | S<0.5 | S | S<4 | S<1 | S<1 | S<0.5 | S<32 | S<0.25 | R>8 | S<1 | S<8/4 | S<0.5/9.5 | R>2 | S<4 | S<0.25 |
CL5 | S<4/2 | R>8 | S<1 | S<0.5 | S<8 | S<0.5 | S<0.5 | S | S<4 | S<1 | S<1 | S<0.5 | S<32 | S<0.25 | R>8 | S<1 | S<8/4 | S<0.5/9.5 | R>2 | S<4 | S<0.25 |
CL7 | S<4/2 | R>8 | S<1 | S<0.5 | S<8 | S<0.5 | S<0.5 | S | S<4 | S<1 | S<1 | S<0.5 | S<32 | S<0.25 | R>8 | S<1 | S<8/4 | S<0.5/9.5 | R>2 | S<4 | S<0.25 |
CL8 | S<4/2 | R>8 | S<1 | S<0.5 | S<8 | S<0.5 | S<0.5 | S | S<4 | S<1 | S<1 | S<0.5 | S<32 | S<0.25 | R>8 | S<1 | S<8/4 | S<0.5/9.5 | R>2 | S<4 | S<0.25 |
CS1 | S<4/2 | R>8 | S<1 | S<0.5 | S<8 | S<0.5 | S<0.5 | S | S<4 | S<1 | S<1 | S<0.5 | S<32 | S<0.25 | R>8 | S<1 | S<8/4 | S<0.5/9.5 | R>2 | S<4 | S<0.25 |
CS2 | S<4/2 | R>8 | S<1 | S<0.5 | S<8 | S<0.5 | S<0.5 | S | S<4 | S<1 | S<1 | S<0.5 | S<32 | S<0.25 | R>8 | S<1 | S<8/4 | S<0.5/9.5 | R>2 | S<4 | S<0.25 |
CS3 | S<4/2 | R>8 | S<1 | S<0.5 | S<8 | S<0.5 | S<0.5 | S | S<4 | S<1 | S<1 | S<0.5 | S<32 | S<0.25 | R>8 | S<1 | S<8/4 | S<0.5/9.5 | R>2 | S<4 | S<0.25 |
CS4 | S<4/2 | R>8 | S<1 | S<0.5 | S<8 | S<0.5 | S<0.5 | S | S<4 | S<1 | S<1 | S<0.5 | S<32 | S<0.25 | R>8 | S<1 | S<8/4 | S<0.5/9.5 | R>2 | S<4 | S<0.25 |
MR1 | S<4/2 | S<2 | S<1 | S<0.5 | S<8 | S<0.5 | S<0.5 | S | S<4 | S<1 | S<1 | S<0.5 | S<32 | S<0.25 | S<0.03 | S<1 | S<8/4 | S<0.5/9.5 | S<0.5 | S<4 | S<0.25 |
MS1 | S<4/2 | R>8 | S<1 | S<0.5 | S<8 | R>4 | S<0.5 | S | S<4 | S<1 | S<1 | S<0.5 | S<32 | S<0.25 | R>8 | S<1 | S<8/4 | S<0.5/9.5 | R>2 | S<4 | S<0.25 |
RpCR1 | R>4/2 | R>8 | S<1 | R>4 | S<8 | R>4 | S<0.5 | S | S<4 | S<1 | S<1 | S<0.5 | S<32 | R>2 | R>8 | S<1 | R>16/8 | R>2/38 | R>2 | S<4 | S<0.25 |
SsCR4 | S<4/2 | R>8 | S<1 | S<0.5 | S<8 | S<0.5 | R>4 | S | S<4 | S<1 | R>4 | S<0.5 | S<32 | S<0.25 | R>8 | S<1 | S<8/4 | R>2/38 | S<0.5 | S<4 | S<0.25 |
SwCR2 | S<4/2 | R>8 | S<1 | S<0.5 | S<8 | R>4 | R>4 | S | R>8 | S<1 | S<1 | S<0.5 | S<32 | S<0.25 | R>8 | S<1 | S<8/4 | S<0.5/9.5 | S<0.5 | S<4 | S<0.25 |
The letter R refers to resistance, and S indicates sensitivity. Ba = Bacillus aryabhattai, BaMA26 = B. amyloliquefaciens, BsTA16 = Bacillus subtilis, BsiDA2 = Bacillus siamensis, BtMB9 = B. thuringiensis, Bv = Bacillus velezensis, Rp = Rhizobium pakistanense, Ss = Staphylococcus saccharolyticus, Sw = Staphylococcus warneri. Amoxicillin-clavulanate = AMC, Ampicillin = AMP, Ciprofloxacin = CIP, Clindamycin = CLI, Ceftriaxone = CRO, Daptomycin = DAP, Erythromycin = ERY, Fosfomycin = FOF; Gentamicin = GEN, Levofloxacin = LVX, Linezolid = LZD, Moxifloxacin = MXF, Nitrofurantoin = NIT, Oxacillin = OXA, Penicillin = PEN, Rifampin = RIF, Ampicillin-sulbactam = SAM, Trimetoprim-sulfametoxazol = SXT, Synercid = SYN, Tetracycline = TET, Vancomycin = VAN. M.I.C = Minimum inhibitory concentration.
CONCLUSIONS
The larvae of Galleria mellonella were used in this study as a model insect due to their availability, low cost, basic requirements, and fast results. It was demonstrated that γ-hemolytic bacteria can be pathogenic to humans. The Galleria mellonella test also revealed that the blood hemolysis test is not the only feature needed for inspecting human bacterial virulence. Pathogenic bacteria can possess different virulence factors, but no hemolysin production, and still can cause human diseases. Some strains studied here were resistant to most the antibiotics tested, highlighting the possibility of encountering multi-drug resistance bacterial control agents, which can be also of concern for agricultural use. This work highlights the need of performing several virulence tests, as well as antibiotic resistance assays, additional in vitro tests, genetic testing, molecular analysis and alternative in vivo models before selecting bacterial strains for agricultural purposes, especially if they reach consumers.
ACKNOWLEDGMENT
M.M.R. Khalil acknowledges the Consejo Nacional de Ciencia y Tecnología (CONACyT) of Mexico and the BEIFI program from Instituto Politécnico Nacional (IPN) for the Ph.D. fellowships.
FUNDING
This study was financially supported by grants SIP-20170939, SIP-20181778, SIP 2019–6353, and SIP 2020–1685 from the Instituto Politécnico Nacional (Mexico) and CONACyT (Programa de Estímulos a la Innovación, Proinnova, Grant No. 230186).
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
AUTHORS' CONTRIBUTIONS
Md. Masudur Rahman Khalil, Gloria Margarita Zamudio-Aguilasocho, and Alejandro Miguel Figueroa-López performed all experiments and analyzed the data. M.M.R. Khalil prepared the manuscript with the assistance of all the authors. I.E. Maldonado-Mendoza inspected this study and oversaw the manuscript preparation and funding acquisition. R. Félix-Gastélum participated in part of the study design and, with Maldonado-Mendoza, participated in manuscript corrections and final editing.
REFERENCES
Agata, N., Ohta, M., Arakawa, Y., & Mori, M. 1995. The bceT gene of Bacillus cereus encodes an enterotoxic protein. In Microbiology. Microbiology Society. DOI:10.1099/13500872-141-4-983.
Ahle, C. M., Stødkilde, K., Afshar, M., Poehlein, A., Ogilvie, L. A., Söderquist, B., Hüpeden, J., & Brüggemann, H. 2020. Staphylococcus saccharolyticus: An overlooked human skin colonizer. Microorganisms. DOI:10.3390/microorganisms8081105.
Anduro-Jordan, J. A., Maldonado-Mendoza, I. E., Figueroa-López, A. M., Torres-Vitela, M. del R., Campas-Baypoli, O. N., Sánchez-Machado, D. I., & Cantú-Soto, E. U. 2022. STEC non-0157 strains in meat from Southern Sonora, Mexico and its antibiotic resistance. Veterinaria México OA. DOI:10.22201/fmvz.24486760e.2022.811.
Aujoulat, F., Marchandin, H., Zorgniotti, I., Masnou, A., & Jumas-Bilak, E. 2015. Rhizobium pusense is the main human pathogen in the genus Agrobacterium/Rhizobium. Clinical Microbiology and Infection : The Official Publication of the European Society of Clinical Microbiology and Infectious Diseases. DOI:10.1016/j.cmi.2014.12.005.
Baek, I., Lee, K., Goodfellow, M., & Chun, J. 2019. Comparative genomic and phylogenomic analyses clarify relationships within and between Bacillus cereus and Bacillus thuringiensis: Proposal for the recognition of two Bacillus thuringiensis genomovars. Frontiers in Microbiology. DOI:10.3389/fmicb.2019.01978.
Beeton, M. L., Alves, D. R., Enright, M. C., & Jenkins, A. T. A. 2015. Assessing phage therapy against Pseudomonas aeruginosa using a Galleria mellonella infection model. International Journal of Antimicrobial Agents. DOI:10.1016/j.ijantimicag.2015.04.005.
Bello-López, E., Rocha-Gracia, R. del C., Castro-Jaimes, S., Cevallos, M. Á., Vargas-Cruz, M., Verdugo-Yocupicio, R., Sáenz, Y., Torres, C., Gutiérrez-Cázarez, Z., Arenas-Hernández, M. M. de la P., & Lozano-Zarain, P. 2020. Antibiotic resistance mechanisms in Acinetobacter spp. strains isolated from patients in a paediatric hospital in Mexico. Journal of Global Antimicrobial Resistance. DOI:10.1016/j.jgar.2020.08.014.
Bokhari, H., Ali, A., Noreen, Z., Thomson, N., & Wren, B. W. 2017. Galleria mellonella is low cost and suitable surrogate host for studying virulence of human pathogenic Vibrio cholerae. Gene. DOI: 10.1016/j.gene.2017.07.019.
Browne, N., Heelan, M., & Kavanagh, K. 2013. An analysis of the structural and functional similarities of insect hemocytes and mammalian phagocytes. Virulence. DOI:10.4161/viru.25906.
Brüggemann, H., Poehlein, A., Brzuszkiewicz, E., Scavenius, C., Enghild, J. J., Al-Zeer, M. A., Brinkmann, V., Jensen, A., & Söderquist, B. 2019. Staphylococcus saccharolyticus isolated from blood cultures and prosthetic joint infections exhibits excessive genome decay. Frontiers in Microbiology. DOI:10.3389/fmicb.2019.00478.
Chen, S., Liu, J., Luo, S., Xing, L., Li, W., & Gong, L. 2024. The effects of Bacillus amyloliquefaciens SC06 on behavior and brain function in broilers infected by Clostridium perfringens. Animals. DOI:10.3390/ani14111547.
Chen, Y., Yan, F., Chai, Y., Liu, H., Kolter, R., Losick, R., & Guo, J. 2013. Biocontrol of tomato wilt disease by Bacillus subtilis isolates from natural environments depends on conserved genes mediating biofilm formation. Environmental Microbiology. DOI:10.1111/j.1462-2920.2012.02860.x.
Cordero-Ramírez, J. D., López-Rivera, R., Figueroa-Lopez, A. M., Mancera-López, M. E., Martínez-Álvarez, J. C., Apodaca-Sánchez, M. Á., & Maldonado-Mendoza, I. E. 2013. Native soil bacteria isolates in Mexico exhibit a promising antagonistic effect against Fusarium oxysporum f. Sp. Radicis-lycopersici. Journal of Basic Microbiology. DOI:10.1002/jobm.201200128.
CLSI. 2024. Clinical & Laboratory Standards Institute: CLSI Guidelines; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2024.
Desbois, A. P., & Coote, P. J. 2012. Chapter 2—Utility of greater wax moth larva (Galleria mellonella) for evaluating the toxicity and efficacy of new antimicrobial agents. En A. I. Laskin, S. Sariaslani, & G. M. Gadd (Eds.), Advances in Applied Microbiology. Academic Press. DOI:10.1016/B978-0-12-394805-2.00002-6.
Divyakolu, S., Chikkala, R., Ratnakar, K. S., & Sritharan, V. 2019. Hemolysins of Staphylococcus aureus—An update on their biology, role in pathogenesis and as targets for anti-virulence therapy. Advances in Infectious Diseases. DOI: 10.4236/aid.2019.92007.
Ehling-Schulz, M., Fricker, M., & Scherer, S. 2004. Bacillus cereus, the causative agent of an emetic type of food-borne illness. Molecular Nutrition & Food Research. DOI:10.1002/mnfr.200400055.
Escamilla-Montes, R., Luna-González, A., Flores-Miranda, Ma. del C., Álvarez-Ruiz, P., Fierro-Coronado, J. A., Sánchez-Ortiz, A. C., & Ávila-Leal, J. 2015. Isolation and characterization of potential probiotic bacteria suitable for mollusk larvae cultures. The Thai Journal of Veterinary Medicine, 45(1), 11-21. DOI:10.56808/2985-1130.2614.
Fallon, J., Kelly, J., & Kavanagh, K. 2012. Galleria mellonella as a model for fungal pathogenicity testing. En A. C. Brand & D. M. MacCallum (Eds.), Host-Fungus Interactions: Methods and Protocols. Humana Press. DOI:10.1007/978-1-61779-539-8_33.
Fedhila, S., Daou, N., Lereclus, D., & Nielsen-LeRoux, C. 2006. Identification of Bacillus cereus internalin and other candidate virulence genes specifically induced during oral infection in insects. Molecular Microbiology. DOI:10.1111/j.1365-2958.2006.05362.x.
Fuchs, B. B., & Mylonakis, E. 2006. Using non-mammalian hosts to study fungal virulence and host defense. Host microbe interactions: fungi/Host microbe interactions: parasites/Host microbe interactions: viruses. DOI:10.1016/j.mib.2006.06.004.
Gao, T., Ding, Y., Wu, Q., Wang, J., Zhang, J., Yu, S., Yu, P., Liu, C., Kong, L., Feng, Z., Chen, M., Wu, S., Zeng, H., & Wu, H. 2018. Prevalence, virulence genes, antimicrobial susceptibility, and genetic diversity of Bacillus cereus isolated from pasteurized milk in China. Frontiers in Microbiology. DOI:10.3389/fmicb.2018.00533.
Gera, K., & McIver, K. S. 2013. Laboratory growth and maintenance of Streptococcus pyogenes (The Group A Streptococcus, GAS). Current Protocols in Microbiology. DOI:10.1002/9780471729259.mc09d02s30.
Giannouli, M., Antunes, L. C., Marchetti, V., Triassi, M., Visca, P., & Zarrilli, R. 2013. Virulence-related traits of epidemic Acinetobacter baumannii strains belonging to the international clonal lineages I-III and to the emerging genotypes ST25 and ST78. BMC Infectious Diseases. DOI:10.1186/1471-2334-13-282.
Godreuil, S., Jean-Pierre, H., Morel, J., Darbas, H., Jumas-Bilak, E., Bañuls, A.-L., & Marchandin, H. 2005. Unusual case of spondylodiscitis due to Staphylococcus saccharolyticus. Joint Bone Spine. DOI:10.1016/j.jbspin.2004.08.010.
Granum, P. E. 1994. Bacillus cereus and its toxins. Journal of Applied Bacteriology. DOI:10.1111/j.1365-2672.1994.tb04358.x.
Guinebretière, M.-H., Broussolle, V., & Nguyen, T. C. 2002. Enterotoxigenic profiles of food-poisoning and food-borne Bacillus cereus strains. Journal of Clinical Microbiology. DOI:10.1128/jcm.40.8.3053-3056.2002.
Hanselman, B. A., Kruth, S. A., Rousseau, J., & Weese, J. S. 2009. Coagulase positive staphylococcal colonization of humans and their household pets. The Canadian Veterinary Journal. DOI:PMC2726022/pdf/cvj_09_954.pdf.
Jang, I.-A., Kim, J., & Park, W. 2016. Endogenous hydrogen peroxide increases biofilm formation by inducing exopolysaccharide production in Acinetobacter oleivorans DR1. Scientific Reports. DOI:10.1038/srep21121.
Jorjão, A. L., Oliveira, L. D., Scorzoni, L., Figueiredo-Godoi, L. M. A., Cristina A. Prata, M., Jorge, A. O. C., & Junqueira, J. C. 2018. From moths to caterpillars: Ideal conditions for Galleria mellonella rearing for in vivo microbiological studies. Virulence. DOI:10.1080/21505594.2017.1397871.
Kavanagh, K., & Fallon, J. P. 2010. Galleria mellonella larvae as models for studying fungal virulence. Fungal Biology Reviews. DOI:10.1016/j.fbr.2010.04.001.
Kay, S., Edwards, J., Brown, J., & Dixon, R. 2019. Galleria mellonella infection model identifies both high and low lethality of Clostridium perfringens toxigenic strains and their response to antimicrobials. Frontiers in Microbiology. DOI:10.3389/fmicb.2019.01281.
Khalid, R., Zhang, Y. J., Ali, S., Sui, X. H., Zhang, X. X., Amara, U., Chen, W. X., & Hayat, R. 2015. Rhizobium pakistanensis sp. Nov., isolated from groundnut (Arachis hypogaea) nodules grown in rainfed Pothwar, Pakistan. Antonie van Leeuwenhoek. DOI:10.1007/s10482-014-0326-x.
Khalil, M. R., Fierro-Coronado, R. A., Peñuelas-Rubio, O., Villa-Lerma, A. G., Plascencia-Jatomea, R., Félix-Gastélum, R., & Maldonado-Mendoza, I. E. 2021. Rhizospheric bacteria as potential biocontrol agents against Fusarium wilt and crown and root rot diseases in tomato. Saudi Journal of Biological Sciences. DOI:10.1016/j.sjbs.2021.08.043.
Khalil, U., Younus, M., Asghar, N., Siddiqui, F., Gómez-Duarte, O. G., Wren, B. W., & Bokhari, H. 2016. Phenotypic and genotypic characterization of enteroaggregative Escherichia coli isolates from pediatric population in Pakistan. APMIS. DOI:10.1111/apm.12577.
Kim, J.-S., Yoon, S.-J., Park, Y.-J., Kim, S.-Y., & Ryu, C.-M. 2020. Crossing the kingdom border: Human diseases caused by plant pathogens. Environmental Microbiology. DOI:10.1111/1462-2920.15028.
Kuchibiro, T., Hirayama, K., Houdai, K., Nakamura, T., Ohnuma, K., Tomida, J., & Kawamura, Y. 2018. First case report of sepsis caused by Rhizobium pusense in Japan. En JMM Case Reports. Microbiology Society. DOI:10.1099/jmmcr.0.005135.
Legius, B., Landuyt, K. V., Verschueren, P., & Westhovens, R. 2012. Septic arthritis due to Staphylococcus warneri: A diagnostic challenge. DOI:10.2174/1874312901206010310.
Lehmann, V. 1973. The nature of phospholipase C from Acinetobacter calcoaceticus: effects on whole red cells and red cell membranes. Acta Pathologica Microbiologica Scandinavica Section B Microbiology and Immunology. DOI:10.1111/j.1699-0463.1973.tb02225.x
Leitão, J. H. 2020. Microbial virulence factors. International Journal of Molecular Sciences. DOI:10.3390/ijms21155320
Lereclus D, Agaisse H, Gominet M, Salamitou S, & Sanchis V. 1996. Identification of a Bacillus thuringiensis gene that positively regulates transcription of the phosphatidylinositol-specific phospholipase C gene at the onset of the stationary phase. Journal of Bacteriology. DOI:10.1128/jb.178.10.2749-2756.1996.
Liu, C.-J., Sun, B., Guo, J., He, J.-L., Feng, B., Wang, H.-G., Cao, K.-K., Liu, T., & Shen, D.-X. 2015. A case of bone marrow infection by Staphylococcus saccharolyticus. European Review for Medical and Pharmacological Sciences. DOI:https://www.europeanreview.org/article/8733
Lund, T., De Buyser, M.-L., & Granum, P. E. 2000. A new cytotoxin from Bacillus cereus that may cause necrotic enteritis. Molecular Microbiology. DOI:10.1046/j.1365-2958.2000.02147.x.
Mancilla-Rojano, J., Ochoa, S. A., Reyes-Grajeda, J. P., Flores, V., Medina-Contreras, O., Espinosa-Mazariego, K., Parra-Ortega, et al. 2020. Molecular epidemiology of Acinetobacter calcoaceticus-Acinetobacter baumannii complex isolated from children at the hospital infantil de Héxico Federico Gómez. Frontiers in Microbiology. DOI:10.3389/fmicb.2020.576673
Matereke, L. T., & Okoh, A. I. 2020. Listeria monocytogenes virulence, antimicrobial resistance and environmental persistence: A review. Pathogens. DOI:10.3390/pathogens9070528
McMillan, S., Verner-Jeffreys, D., Weeks, J., Austin, B., & Desbois, A. P. 2015. Larva of the greater wax moth, Galleria mellonella, is a suitable alternative host for studying virulence of fish pathogenic Vibrio anguillarum. BMC Microbiology. DOI:10.1186/s12866-015-0466-9
Ménard, G., Rouillon, A., Cattoir, V., Donnio, P. 2021. Galleria mellonella as a suitable model of bacterial infection: past, present and future. Frontiers in Cellular and Infection Microbiology. DOI: 10.3389/fcimb.2021.782733.
Méndez, I., Fallard, A., Soto, I., Tortella, G., de la Luz Mora, M., Valentine, A. J., Barra, P. J., & Duran, P. 2021. Efficient biocontrol of Gaeumannomyces graminis var. Tritici in wheat: Using bacteria isolated from suppressive soils. Agronomy. DOI:10.3390/agronomy11102008
Meng, L., Liu, H., Lan, T., Dong, L., Hu, H., Zhao, S., Zhang, Y., Zheng, N., & Wang, J. 2020. Antibiotic resistance patterns of Pseudomonas spp. isolated from raw milk revealed by whole genome sequencing. Frontiers in Microbiology. DOI:10.3389/fmicb.2020.01005
Mikhael, M. M., Bach, H. G., Huddleston, P. M., Maus, T. P., & Berbari, E. F. 2009. Multilevel diskitis and vertebral osteomyelitis after diskography. Orthopedics. DOI:10.3928/01477447-20090101-16
Misawa, N., Hirayama K, Itoh K, & Takahashi E. 1995. Detection of alpha- and beta-hemolytic-like activity from Campylobacter jejuni. Journal of Clinical Microbiology. DOI:10.1128/jcm.33.3.729-731.1995
Morin, C. D., Déziel, E., Gauthier, J., Levesque, R. C., & Lau, G. W. 2021. An organ system-based synopsis of Pseudomonas aeruginosa virulence. Virulence. DOI:10.1080/21505594.2021.1926408
Nurjadi, D., & Boutin, S. (2022). Acinetobacter baumannii. En I. de Filippis (Ed.), Molecular Typing in Bacterial Infections, Volume II. Springer International Publishing. DOI:10.1007/978-3-030-83217-9_6.
Pakbin, B., Brück, W. M., & Rossen, J. W. A. 2021. Virulence factors of enteric pathogenic Escherichia coli: A review. International Journal of Molecular Sciences. DOI:10.3390/ijms22189922.
Peleg, A. Y., de Breij, A., Adams, M. D., Cerqueira, G. M., Mocali, S., Galardini, M., Nibbering, P. H., Earl, A. M., Ward, D. V., Paterson, D. L., Seifert, H., & Dijkshoorn, L. 2012. The success of Acinetobacter species; genetic, metabolic and virulence attributes. PLOS ONE. DOI:10.1371/journal.pone.0046984.
Peleg, A. Y., Jara, S., Monga Divya, Eliopoulos George M., Moellering Robert C., & Mylonakis Eleftherios 2009. Galleria mellonella as a model system to study Acinetobacter baumannii pathogenesis and therapeutics. Antimicrobial Agents and Chemotherapy. DOI:10.1128/aac.01533-08
Poria, V., Dębiec-Andrzejewska, K., Fiodor, A., Lyzohub, M., Ajijah, N., Singh, S., & Pranaw, K. 2022. Plant Growth-Promoting Bacteria (PGPB) integrated phytotechnology: A sustainable approach for remediation of marginal lands. Frontiers in Plant Science. DOI:10.3389/fpls.2022.999866.
Quereda, J. J., Andersson, C., Cossart, P., Johansson, J., & Pizarro-Cerdá, J. 2018. Role in virulence of phospholipases, listeriolysin O and listeriolysin S from epidemic Listeria monocytogenes using the chicken embryo infection model. Veterinary Research. DOI:10.1186/s13567-017-0496-4.
Rahimi, A., Siavash Moghaddam, S., Ghiyasi, M., Heydarzadeh, S., Ghazizadeh, K., & Popović-Djordjević, J. 2019. The influence of chemical, organic and biological fertilizers on agrobiological and antioxidant properties of Syrian cephalaria (Cephalaria Syriaca L.). Agriculture. DOI:10.3390/agriculture9060122.
Ramarao, N., Nielsen-Leroux, C., & Lereclus, D. 2012. The insect Galleria mellonella as a powerful infection model to investigate bacterial pathogenesis. JoVE. DOI:10.3791/4392.
Ravi Patel, D. N. B. 2023. Isolation and characterization of robust plant growth-promoting rhizobacteria from lignite mines, Gujarat. En Journal of Applied Biology & Biotechnology. DOI:10.7324/JABB.2023.110225.
Rivas-Ruiz, C. M., Cantú-Soto, E. U., Maldonado-Mendoza, I. E., Figueroa-López, A. M., Anduro-Jordan, J. A., Luna-Nevarez, P., & López-Castro, P. A. 2020. Detección de Escherichia coli productora de toxina-Shiga en bovinos asintomáticos del sur de Sonora, México. Ecosistemas y Recursos Agropecuarios. DOI:10.19136/era.a7n2.2240
Salgado-Morales, R., Martínez-Ocampo, F., Obregón-Barboza, V., Vilchis-Martínez, K., Jiménez-Pérez, A., & Dantán-González, E. 2019. Assessing the pathogenicity of two bacteria isolated from the entomopathogenic nematode heterorhabditis indica against Galleria mellonella and some pest insects. Insects. DOI:10.3390/insects10030083.
Schreiter, S., Babin, D., Smalla, K., & Grosch, R. 2018. Rhizosphere competence and biocontrol effect of Pseudomonas sp. RU47 independent from plant species and soil type at the field scale. Frontiers in Microbiology. DOI:10.3389/fmicb.2018.00097.
Senior, N. J., Bagnall, M. C., Champion, O. L., Reynolds, S. E., La Ragione, R. M., Woodward, M. J., Salguero, F. J., & Titball, R. W. 2011. Galleria mellonella as an infection model for Campylobacter jejuni virulence. En Journal of Medical Microbiology. Microbiology Society. DOI:10.1099/jmm.0.026658-0
Smith, D. J., Ramsay, K. A., Yerkovich, S. T., Reid, D. W., Wainwright, C. E., Grimwood, K., Bell, S. C., & Kidd, T. J. 2016. Pseudomonas aeruginosa antibiotic resistance in Australian cystic fibrosis centres. Respirology. DOI:10.1111/resp.12714.
Strateva, T., & Mitov, I. 2011. Contribution of an arsenal of virulence factors to pathogenesis of Pseudomonas aeruginosa infections. Annals of Microbiology. DOI:10.1007/s13213-011-0273-y
Sunenshine, R. H., Wright, M.-O., Maragakis, L. L., Harris, A. D., Song, X., Hebden, J., et al. 2007. Multidrug-resistant Acinetobacter infection mortality rate and length of hospitalization. Emerging Infectious Disease Journal.DOI:10.3201/eid1301.060716.
Trojani, M.-C., Lamy, B., Ruimy, R., Amoretti, N., Risso, K., & Roux, C. 2020. An unusual Staphylococcus saccharolyticus spondylodiscitis post kyphoplasty: A case report. BMC Infectious Diseases. DOI:10.1186/s12879-020-05263-5
Tsai, C. J.-Y., Loh, J. M. S., & Proft, T. 2016. Galleria mellonella infection models for the study of bacterial diseases and for antimicrobial drug testing. Virulence. DOI:10.1080/21505594.2015.1135289.
Vera, A., González, G., Domínguez, M., & Bello, H. 2013. Principales factores de virulencia de Listeria monocytogenes y su regulación. Revista chilena de infectología. DOI:10.4067/S0716-10182013000400010.
Vidaillac, C., & Chotirmall, S. H. 2021. Pseudomonas aeruginosa in bronchiectasis: Infection, inflammation, and therapies. Expert Review of Respiratory Medicine. DOI:10.1080/17476348.2021.1906225.
Wagley, S., Borne, R., Harrison, J., Baker-Austin, C., Ottaviani, D., Leoni, F., Vuddhakul, V., & Titball, R. W. 2018. Galleria mellonella as an infection model to investigate virulence of Vibrio parahaemolyticus. Virulence. DOI:10.1080/21505594.2017.1384895.
Wang, P., Liu, Y., Xu, Y., & Xu, Z. 2020. Staphylococcus saccharolyticus infection: Case series with a PRISMA-compliant systemic review. Medicine. DOI: 10.1097/MD.0000000000020686.
Wang, X., Zhou, X., Cai, Z., Guo, L., Chen, X., Chen, X., Liu, J., Feng, M., Qiu, Y., Zhang, Y., & Wang, A. 2021. A biocontrol strain of Pseudomonas aeruginosa CQ-40 promote growth and control Botrytis cinerea in tomato. Pathogens. DOI:10.3390/pathogens10010022.
Wertheim, H. F., Melles, D. C., Vos, M. C., van Leeuwen, W., van Belkum, A., Verbrugh, H. A., & Nouwen, J. L. 2005. The role of nasal carriage in Staphylococcus aureus infections. The Lancet Infectious Diseases. DOI:10.1016/S1473-3099(05)70295-4.
Westblom, T. U., Gorse, G. J., Milligan, T. W., & Schindzielorz, A. H. 1990. Anaerobic endocarditis caused by Staphylococcus saccharolyticus. Journal of Clinical Microbiology. DOI:10.1128/jcm.28.12.2818-2819.1990.
WHO. 2020. Worlth Health Organization, Geveva. Antibiotic resistance. Revisado en:www.who.int/news-room/fact-sheets/detail/antibiotic-resistance
Wolfmeier, H., Wardell, S. J. T., Liu, L. T., Falsafi, R., Draeger, A., Babiychuk, E. B., Pletzer, D., & Hancock, R. E. W. 2022. Targeting the Pseudomonas aeruginosa virulence factor phospholipase C with engineered liposomes. Frontiers in Microbiology. DOI:10.3389/fmicb.2022.867449.
Wu, X., Yu, C., & Wang, X. 2009. A case of Staphylococcus saccharolyticus pneumonia. International Journal of Infectious Diseases. DOI:10.1016/j.ijid.2008.06.007.
Young, N., & Bhally, H. 2017. Bilateral neck pyomyositis caused by Staphylococcus capitis and Staphylococcus saccharolyticus in a diabetic adult. Case Reports in Infectious Diseases. DOI:10.1155/2017/3713212.
Zamudio-Aguilasocho, G. M. Selección y evaluación de bacterias endófitas de plantas de la familia Poaceae de la Reserva la Uba, Guasave, Sinaloa, con potencial biotecnológico y antagónico a Rhizoctonia zeae. [Tesis de Posgrado] Guasave: Instituto Politécnico Nacional; 2019.
Bioc Scientia 2026 Acceso libre
[a]Ajusta la fuente a Times new roman, numero 10 intr. Sencillo para todo el texto