Camacho-Haro et al., 2025 de
Artículo de revisión
Interacción de los exudados radicales con nematodos del género
Meloidogyne y otros microorganismos en la rizosfera
Camacho-Haro Marcos1, García-Ulloa Manuel1, Camacho-Báez Jesús Ricardo1, Montiel-Montoya Jorge1, Magallanes-Tapia Marco Antonio1.
1Instituto Politécnico Nacional. Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional, Unidad Sinaloa. Juan de Dios Bátiz Paredes 250. Col San Joachín. CP 81101 Guasave, Sinaloa, México
Correspondencia: mmagallanes@ipn.mx
Área Temática: Ciencias de las Plantas Recibido: 27 junio, 2025 Aceptado:08 septiembre, 2025 Publicado: 17 septiembre 2025 Cita: Camacho-Haro M, García-Ulloa M, Camacho-Báez JR, Montiel-Montoya J, y Magallanes-Tapia MA. 2025. Interacción de los exudados radicales con nematodos del género Meloidogyne y otros microorganismos en la rizosfera. Bioc Scientia 2(1): 2025. https://doi.org/10.63622/RBS.2515 Copyright: © 2024 by the authors. Submitted for possible open access publication under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by-nc/4.0/). |
Resumen: Las plantas establecen interacciones fundamentales con una amplia diversidad de organismos, lo que cobra especial relevancia con sistemas de producción hortícola. En la rizosfera, las raíces generan un escenario altamente dinámico y vital, donde coexisten microorganismos benéficos y patógenos. En esta interacción, el Phylum Nemata destaca por su predominio en agroecosistemas, cuyas especies interactúan con las raíces de las plantas. El género Meloidogyne representa uno de los grupos de nematodos fitoparásitos más dañinos y económicamente el más importante en el mundo, responsable de millonarias pérdidas en cultivos hortícolas. Como respuesta defensiva, las plantas han desarrollado estrategias radicales basadas en su metabolismo secundario que sintetizan una variedad de compuestos bioactivos incluidos los semioquímicos que participan en la protección contra patógenos. Entre estos, destacan los flavonoides, que son moléculas biológicamente activas con una amplia gama de actividades y diversidad estructural, particularmente asociadas a la fisiología, crecimiento y desarrollo de las plantas. No obstante, el conocimiento actual sobre cómo aprovechar este potencial defensivo de los metabolitos secundarios para el manejo eficaz de estos fitoparásitos sigue siendo limitado. La presente revisión aborda información clave sobre las interacciones planta-microorganismo en la rizosfera, enfatizando la relevancia de estos compuestos en el manejo de plagas y enfermedades; así como la mejora de la planta. De igual manera, se señalan áreas críticas donde se requiere mayor investigación, con miras a desarrollar estrategias de manejo más eficientes y sostenibles dentro de la horticultura.
Palabras clave: Metabolitos secundarios, semioquímicos, flavonoides, agroecosistemas, ecología
Abstract: Plants establish fundamental interactions with a wide diversity of organisms-particularly relevant in horticultural production systems. Within the rhizosphere, roots create a highly dynamic and vital environment where beneficial and pathogenic microorganisms coexist. Among these biotic agents, nematodes of the phylum Nemata (syn. Nematoda) are predominant in agroecosystems, and root-knot nematodes of the genus Meloidogyne rank among the most damaging and economically significant plant-parasitic nematodes worldwide, causing substantial losses in horticultural crops. As a defensive response, plants deploy root-based strategies grounded in their secondary metabolism, synthesizing and often exuding diverse bioactive compounds, including semiochemicals, that contribute to pathogen protection. Flavonoids are especially noteworthy for their wide structural diversity and functional roles closely linked to plant physiology, growth, and development. Nevertheless, current understanding of how to harness the defensive potential of secondary-metabolite-derived root exudates for effective management of these phytoparasites remains limited. This review synthesizes key information on plant–microorganism interactions in the rhizosphere, emphasizing the relevance of these compounds for pest and disease management as well as crop improvement, and it highlights critical knowledge gaps that must be addressed to develop more efficient and sustainable management strategies in horticulture.
Keywords: Secondary metabolites, semiochemicals, flavonoids, agroecosystems, ecology.
INTRODUCCIÓN
Las plantas, como organismos sésiles, interactúan con una amplia gama de seres vivos, incluidos artrópodos, herbívoros, hongos, bacterias, nematodos, etc., y estas interacciones pueden modificar su fisiología y afectar el comportamiento, rendimiento y abundancia de otras especies circundantes (Hakim et al., 2021; Chamkhi et al., 2022).
En la rizósfera, las raíces participan en múltiples dinámicas como raíz-raíz, raíz-insecto y raíz-patógenos (Barton et al., 2018). Además de estas relaciones biológicas, también establecen procesos químicos, físicos y biológicos con el entorno circundante que se enriquecen con los exudados radicales (Hawes et al., 2016); los cuales, liberan un alto contenido de compuestos químicos que modulan la atracción o repulsión de diversos organismos patógenos o beneficiosos, y pueden ser determinantes en la salud de la planta (Rolfe et al., 2019).
Un caso puntual de señalización radical son las isoflavonas de la soya (Glycine max L.), que atraen tanto a bacterias fijadoras de nitrógeno (Bradyrhizobium japonicum; Castro-Barquero, 2024), como a Phytophthora sojae ─un oomiceto patógeno (López-Cardona y López-Casalla, 2018), lo que evidencia la doble naturaleza de ciertos compuestos químicos. Este tipo de interacciones ilustra como los exudados radicales pueden fomentar relaciones mutualistas o facilitar el ataque de patógenos en la rizósfera (De-la-Peña y Loyola-Vargas, 2014; Zhang et al., 2020).
Esta revisión se centra en evidencia reciente vinculada al eje químico de los exudados radicales. Para ello, se aplicaron los siguientes criterios de inclusión: a) extractos o aceites cuyos principales compuestos (flavonoides, ácidos fenólicos, terpenoides) están documentados también en exudados; b) vínculo explícito con mecanismos plausibles sobre nematodos agalladores (Meloidogyne spp.), ya sea quimiotaxis/atracción-repelencia, ovicidia/nematostasia o inducción de defensas en raíz; y c) ensayos recientes (preferentemente 2020–2025) incluyendo in vitro y maceta/microparcela. Se excluyeron listados enciclopédicos y trabajos sin identificación química mínima o sin resultado biológico verificable.
Metabolismo secundario en plantas (MSP)
El MSP desempeña un papel crucial en la síntesis de compuestos que no solo se protegen contra insectos y enfermedades; sino que también, inhiben el crecimiento de otras especies (Jan et al., 2021). Por su diversidad de propiedades biológicas, estos metabolitos se han utilizado históricamente en medicina y otras aplicaciones (Jamshidi-Kia et al., 2018). Ciertas plantas se caracterizan por su contenido de compuestos fitoquímicos y sustancias inorgánicas, como sales de ácido silícico y potasio, que constituyen más del 10% de su composición. Además, contienen esteroles, flavonoides y varios ácidos (ascórbico, dicarboxílico, fenólico y polienico) que les confieren capacidades antimicrobianas y antiinflamatorias (Más Toro et al., 2017; Sousa et al., 2019).
De igual manera, los aceites esenciales y extractos de plantas son ricos en múltiples sustancias bioactivas como alcaloides, glucósidos cianogénicos, glucosinolatos, lípidos, fenólicos, terpenos y poliacetilenos (Campo Fernández et al., 2018; Macêdo et al., 2020; Tlak Gajger y Dar, 2021). En conjunto, estos metabolitos secundarios (MS) resaltan la complejidad del MSP y su importancia evolutiva en la defensa contra patógenos; así como la tolerancia al estrés ambiental (Ben Mrid et al., 2021; Yadav et al., 2021).
Aplicaciones de metabolitos en defensa y salud de plantas
Algunas proteínas como las reguladoras de información silenciosa (SIR, por sus siglas en inglés) desempeñan un papel importante en el ajuste del tiempo, la ubicación y el nivel de expresión genética en el proceso de defensa de las plantas contra depredadores (insectos) o patógenos. Por ejemplo, Blanco-Labra y Aguirre-Mancilla (2002) destacan inhibidores de enzimas hidrolíticas (proteasas y amilasas) y otras relacionadas con la patogénesis (β-1,3-glucanasas y endoquitinasas), para combatir enfermedades en maíz y tabaco, respectivamente. Menéndez et al. (2020) mencionan que diferentes genes codifican proteínas para diversas funciones: reforzar o reparar la pared celular, participar en procesos antimicrobianos y activarse en presencia de un patógeno. Aunque no son MS per se, estas proteínas pueden incidir en la biosíntesis y liberación de compuestos bioactivos incluidos aquellos que la raíz exuda hacia la rizósfera que las plantas utilizan para su defensa y otras funciones fisiológicas (Martínez-Bastidas et al., 2017).
Esta modulación de perfiles metabólicos y exudados es consistente con efectos reportados sobre la atracción/quimiotaxis, la eclosión y el establecimiento de nematodos fitoparásitos, lo que sugiere su integración en esquemas de manejo sustentable (Ngala et al., 2024; Chen et al., 2025). Asimismo, el desarrollo de métodos que fortalezcan las vías de síntesis en la planta sería altamente relevante para la producción de bioplaguicidas y representa una necesidad urgente (Dassanayake et al., 2021).
Ecología química y manejo de plagas y enfermedades
Semioquímicos y su función
La ecología química estudia la manera en cómo los compuestos químicos intervienen en las interacciones entre los seres vivos, incluidas aquellas entre plantas, animales y el ambiente (Acevedo, 2020). En este contexto, se denomina semioquímicos a las sustancias que un organismo produce para modificar el comportamiento o la fisiología de otros (Ayelo et al., 2022). En su mayoría, estos compuestos son volátiles y participan en procesos tan diversos como la atracción de polinizadores, la defensa contra plagas o la comunicación inter/intraespecífica. Dentro de esta categoría, los compuestos orgánicos volátiles (COV), constituyen un subgrupo particularmente relevante, pues se liberan a temperatura ambiente y pueden tener efectos en la dinámica ecológica (Holopainen et al., 2018; Cantúa Ayala et al., 2019; Meents y Mithöfer, 2020). Sin embargo, pese a los notables avances en la identificación de una amplia diversidad de semioquímicos, la mayoría de los descubrimientos se centra en la parte aérea de la planta, subestimando la complejidad de las interacciones químicas que ocurren en la rizosfera (Korenblum et al., 2022; Schwerdtner y Spohn, 2022).
Bajo condiciones de estrés, ya sea biótico (por ataque de plagas o patógenos) o abiótico (cambios de temperatura, radiación, etc.), diversas plantas liberan mezclas de COV, como terpenos, derivados de ácidos grasos y compuestos aromáticos (Ramya et al., 2020; Lu et al., 2022). Estos pueden repeler insectos y nematodos fitoparásitos, actuando además como disparadores de mecanismos de defensa o de atracción de polinizadores (Kirwa et al., 2018; Meents y Mithöfer, 2020). Ciertos estudios sugieren ─incluso─ su potencial aplicación en programas de manejo integrado de plagas y enfermedades (Blassioli-Moraes et al., 2019). No obstante, la recopilación y caracterización de COV en la rizósfera presenta retos metodológicos considerables, por lo que se requieren mejores técnicas para profundizar en el conocimiento sobre su papel en este ambiente (Torto et al., 2018).
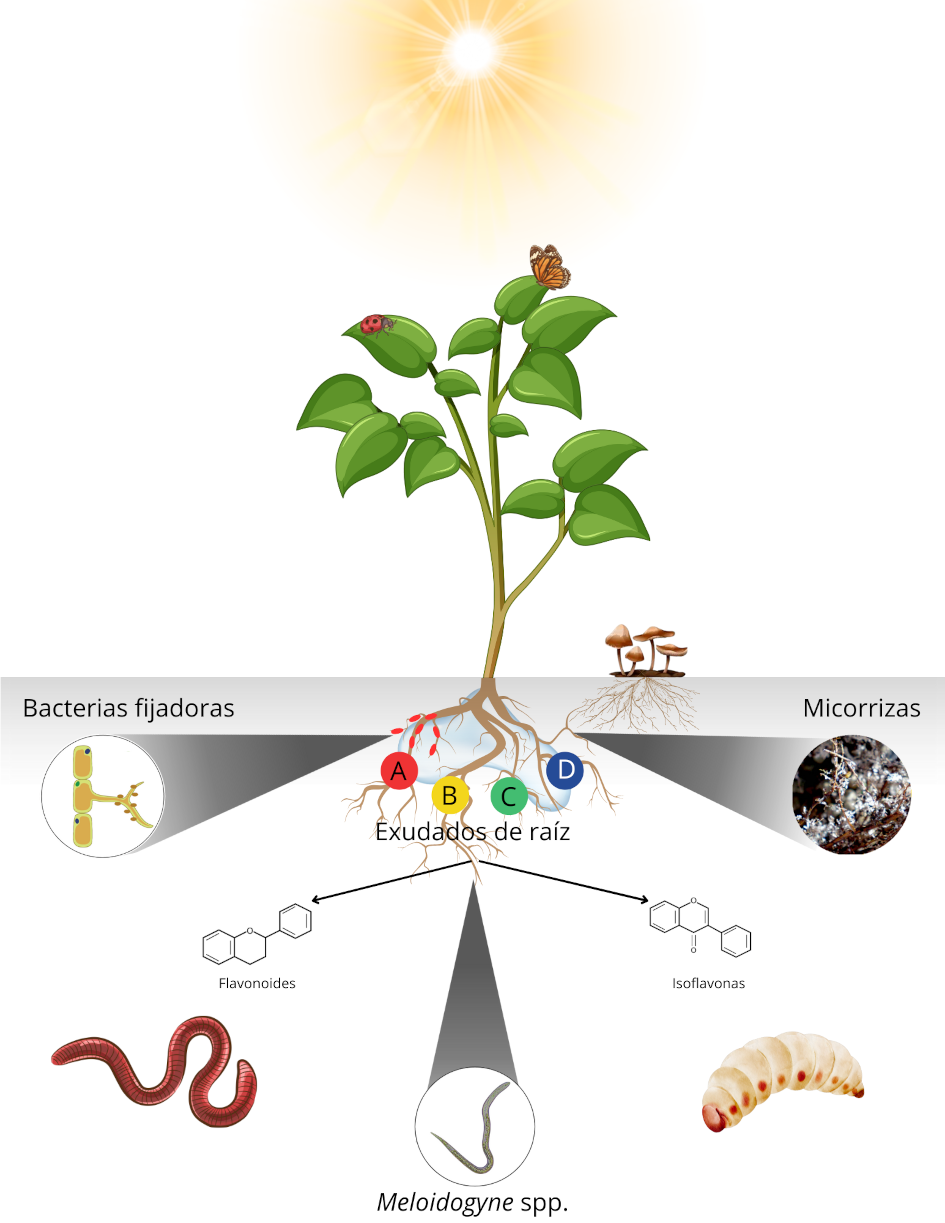
Además, los organismos simbiontes presentes en el suelo pueden influir en la evolución de los mecanismos de defensa vegetal (Figura 1). Un parásito específico no solo modifica la respuesta inmune de la planta, sino también interactúa con seres mutualistas y competidores que coexisten en el mismo nicho. Por ello, la rizósfera se revela como un escenario sumamente dinámico y vital para estudiar la forma en que los semioquímicos ─y en particular los COV─ afectan la salud y productividad de las plantas (Zélé et al., 2018).
Figura 1. Relación integral de los exudados radicales con Meloidogyne spp., microorganismos simbiontes y otras plagas en la rizósfera. A) aceites esenciales, B) sales, C) ácidos y D) proteínas reguladoras de información silenciosa (SIR). |
Nematodos fitoparásitos y la rizósfera
Además de los artrópodos y microorganismos benéficos, las plantas ejercen interacciones edáficas con los nematodos fitoparásitos (Biere y Goverse, 2016; Park y Ryu, 2021). El Phylum Nemata, con 80 000 a 500 000 especies descritas, es el más diverso después de los artrópodos (El-Saadony et al., 2021), y ─en particular─ el género Meloidogyne se considera el grupo de nematodos parásitos de plantas (NPP) más perjudicial y económicamente el de mayor importancia para la agricultura, ya que es capaz de causar pérdidas de hasta 150 mil millones de dólares anuales (Liu y Park, 2018; Oosterbeek et al., 2021; Palomares-Rius et al., 2021). En su etapa infectiva, los NPP se orientan por los exudados de las raíces para ubicar sus sitios de alimentación (Figura 1) (Gautier et al., 2021; Mathesius y Costa, 2021); no obstante, todavía se desconoce el mecanismo sobre como los COV señalan a los nematodos la ubicación de las raíces (Guerrieri et al., 2021).
En tomate (Solanum lycopersicum L.), se documentó que el ácido jasmónico contribuye a la resistencia mediada por el gen Mi-1 contra Meloidogyne, lo que muestra la complejidad de las rutas hormonales involucradas en la defensa de plantas (Seiml-Buchinger et al., 2019). Además, las asociaciones mutualistas en la rizosfera podrían influir en la ecología química de los NPP, y cómo este conocimiento puede utilizarse para su manejo integrado (Ochola et al., 2021).
Interacciones mutualistas y manejo de nematodos
Los organismos mutualistas, como las rizobacterias y los hongos micorrícicos, pueden alterar la concentración de nutrientes y MS en la rizósfera (Hu et al., 2018). Comprender esta comunicación química es fundamental para diseñar estrategias de control, ya sea de forma aislada o como parte de un manejo integrado de NPP (Torto et al., 2018; Ochola et al., 2021). Por ejemplo, los flavonoides se inducen en las raíces durante la infección por NPP, desempeñando múltiples roles en la defensa y señalización; no obstante, se desconocen muchos de sus mecanismos funcionales (Chin et al., 2018; Eugui et al., 2022). También, se ha propuesto incrementar la concentración de flavonoides (que actúan como fitoalexinas), mediante ingeniería genética, con el objetivo de mejorar la resistencia contra NPP (Chin et al., 2018). Aún con ello, ciertos nematodos como Meloidogyne en tomate consiguen establecerse y causar infecciones exitosas, lo que sugiere que sus hábitos de migración intercelular evaden en parte las defensas de la planta (Figura 1) (Holbein et al., 2019). Las fitohormonas también pueden inhibir o modular estas interacciones, pero se desconocen en profundidad sus rutas funcionales y de señalización (Sikder et al., 2021).
Estrategias botánicas para el control de patógenos
Plantas antagonistas y extractos botánicos
En las últimas décadas, se ha observado que los patógenos han desarrollado resistencia a los plaguicidas químicos (Miller et al., 2022), lo que destaca la necesidad de explorar alternativas naturales amigables con el ambiente. En este sentido, los extractos botánicos surgen como una de las alternativas más prometedoras para el control y manejo de patógenos (Lengai et al., 2020; Vélez-Ruíz et al., 2022; Shai et al., 2024). Spiegler et al. (2017) examinaron distintos extractos de plantas con composición polifenólica; entre ellos, taninos condensados e hidrolizables, flavonoides y fenilpropanoides, algunos de los cuales son reconocidos por su acción contra fitopatógenos (Menikheim et al., 2024). Desde la antigüedad, los extractos de plantas y diversos productos naturales se han utilizado como tratamientos complementarios a una variedad de enfermedades y plagas (Gahukar, 2012).
Los compuestos fenólicos comprenden un gran grupo de MS, entre los cuales destacan los flavonoides, proantocianidinas y taninos hidrolizables; así como ésteres depsídicos, ácidos fenólicos, fenilpropanoides, lignanos, estilbenoides, e incluso ciertas xantonas y cumarinas. Aun así, se necesita profundizar en la investigación sobre la actividad de los flavonoides, pues la información aún es limitada en comparación con la generada sobre taninos condensados. Comprender sus mecanismos de acción e interacción con otros componentes de los extractos permitirá aprovechar mejor las plantas antagonistas frente a diversos fitoparásitos, incluidos los NPP (Spiegler et al., 2017).
Aplicación en nematodos agalladores (Meloidogyne spp.)
Diversos extractos botánicos concentran flavonoides, terpenoides y fenoles que las raíces exudan de manera natural; por ello, pueden considerarse análogos funcionales de los exudados radicales y, en este sentido, parte del mismo sistema químico que modula la atracción, la eclosión y el establecimiento de Meloidogyne (Cuadro 1). En años recientes, los extractos polifenólicos y aceites esenciales han mostrado resultados consistentes: en bioensayos in vitro inhiben la eclosión y la movilidad de J2 de M. incognita y, en tomate en maceta o microparcela, reducen significativamente el índice de agallamiento y las poblaciones, con desempeños comparables a nematicidas de referencia (D’Addabbo et al., 2025; Vimala et al., 2025).
Cuadro 1. Extractos o aceites botánicos contra Meloidogyne spp. (2020-2025): evidencia in vitro e in planta.
Material botánico / formulación | Diseño de evaluación | Condición de ensayo / cultivo | Efecto principal vs. Meloidogyne spp. | Referencias |
Aceites esenciales de Mentha spicata y Piper longum + Bacillus spp. con enmiendas orgánicas) | In vitro + microparcela | Tomate (microparcela) | Inhibición de eclosión/movilidad y < agallas | Vimala et al. (2025) |
Extracto etanólico de pericarpios de Citrus microcarpa | In vitro (mortalidad J2, eclosión) | M. enterolobii | 100% mortalidad a 48 h (50 mg mL⁻¹); 89.3% inhibición de eclosión a 24 h | Dou et al. (2024) |
(E)-cinnamaldehído y AE de Cinnamomum cassia | In vitro + maceta | Tomate / papa | > 68% - 90% mortalidad, o < eclosión (según dosis), < infestación en maceta | D’Addabbo et al. (2025) |
AE de Cinnamomum zeylanicum y Citrus aurantium | In vitro | J2 de M. incognita | Alta mortalidad (C. zeylanicum) | Fanelli et al. (2025) |
Isotiocianato de alilo (AITC -compuesto de Brassicaceae) | In vitro + maceta/bandeja; biofumigación | M. hapla | Inmovilización de J2; < de agallas cuando se suplementa AITC a la biofumigación | Dahlin y Hallmann (2020) |
AE de Origanum vulgare subsp. glandulosum | In vitro | M. incognita | Mortalidad y efecto sobre eclosión; perfil fitoquímico caracterizado | Mezerket et al. (2025) |
Mezclas sinérgicas de AE (varias especies) | In vitro + planta | Tomate y otros | Sinergia contra M. incognita y fitopatógenos fúngicos | Bargali et al. (2024) |
Combinación de compuestos (extractos vegetales) | In vitro + maceta | Tomate | < significativa de agallas; inducción de defensas | Degroote et al. (2024) |
Monoterpenos (carvona, cuminaldehído, linalool) | Invernadero (y análisis de defensas) | Tomate | < reproducción/agallas; > expresión de genes de defensa | Elsharkawy et al. (2022) |
Nanoemulsión de AE de Carlina acaulis | In vitro + maceta | Tomate | Eficacia mejorada por nanoformulación; < J2 y agallas | Ntalli et al. (2022) |
Extractos de especias (clavo, canela, etc.) | in vitro + maceta (invernadero) | M. arenaria | < movilidad/mortalidad J2 in vitro; cinco extractos más activos redujeron agallas en maceta (tomate) | Haroun et al. (2022) |
AE de Allium sativum (ajo) | Invernadero | Tomate | < poblaciones de M. incognita y agallas | Jardim et al. (2020) |
Extractos de Maerua angolensis y Tabernaemontana elegans | Invernadero | Tomate | Aplicación antes o al inocular ≈ Nemacur® en reducción poblacional | Mnyambo et al. (2024) |
AE = aceite esencial; J2 = juveniles de 2do estadio
Metabolitos secundarios (MS) de interés
Flavonoides
Son polihidroxifenoles de bajo peso molecular sintetizados por las plantas a través de la vía del ácido shikímico, y cuentan con gran potencial terapéutico, incluyendo su capacidad para combatir diversas infecciones. A pesar de que la palabra "flavonoide" proviene de flavous, que significa amarillo, estos compuestos pueden ser de diversos colores como blanco púrpura, rojo o azul (Hazafa et al., 2020; Kapoor et al., 2021; Shen et al., 2022). Estos MS multifuncionales participan en procesos enzimáticos, coloración de flores, señalización, transporte de auxinas, protección contra rayos UV, regulación hormonal de las plantas y defensa contra factores bióticos y abióticos (Khalid et al., 2019; Wen et al., 2020; Roy et al., 2022). Su transporte hacia la rizosfera puede realizarse a través de la descomposición de las células radicales o mediante transportadores casete de unión (ABC, por sus siglas en inglés) a trifosfato de adenosina (ATP, por sus siglas en inglés) (Biała-Leonhard et al., 2021). Además, están involucrados en la fijación biológica del nitrógeno en los nódulos radicales, resultado de la simbiosis entre leguminosas y bacterias del género Rhizobium. Si bien, se ha documentado la exudación de isoflavonoides como la genisteína a través de transportadores ABC en la membrana plasmática de la raíz de soya, aún se requiere más información para comprender completamente estos mecanismos (Khalid et al., 2019; Biała-Leonhard et al., 2021).
Fitoalexinas
Son MS producidos tras una infección, con el fin de frenar la propagación del patógeno (He et al., 2019; Nguyen et al., 2022). Los elicitores son compuestos naturales que estimulan la síntesis de fitoalexinas y activan varios mecanismos de defensa de la planta. Estos incluyen la Resistencia Sistémica Adquirida, asociada con el ácido salicílico y las proteínas relacionadas con la patogénesis; la Resistencia Sistémica Inducida, activada por rizobacterias saprofitas; y la Resistencia Local Adquirida, que se establece tras la respuesta hipersensible de la planta e induce la producción de fitoalexinas (Thakur et al., 2019).
Aceites esenciales
Se caracterizan por su alta volatilidad y por incluir alcoholes aromáticos, ácidos, ésteres, cetonas y aldehídos (Perczak et al., 2019). Estos compuestos pueden funcionar como protectores contra depredadores o como atrayentes de polinizadores. Los terpenoides y fenilpropanoides son aceites esenciales que se usan ampliamente como moléculas bioactivas en áreas de biología, agronomía, medicina y ciencias farmacéuticas (Davison y Brimble, 2019; Boba et al., 2020; Bavi et al., 2022), y se ha documentado que tienen actividades antihelmínticas o insecticidas. Lo anterior, los sitúa como candidatos para el control de plagas y enfermedades, incluidos los NPP (De Carvalho et al., 2019; Monzote et al., 2021; Verrillo et al., 2021; Da Silva Sá et al., 2023).
Otros metabolitos y cuestiones evolutivas
Las plantas sintetizan MS que, si bien no son esenciales para la supervivencia inmediata, desempeñan roles cruciales en la defensa, la atracción de polinizadores y otros procesos ecológicos. Muchos de estos subproductos se consideraban desecho del metabolismo primario, pero investigaciones sobre ecología química han demostrado su enorme valor biológico (Wink, 2015). A pesar de que se han aislado e identificado alrededor de 200 000 MS, esta cifra es pequeña en comparación con las aproximadamente 391 000 especies de plantas descritas (Kessler y Kalske, 2018). Entre estos compuestos, los terpenoides destacan por su función defensiva en numerosos cultivos de importancia agrícola, como el maíz (Zea mays), donde se requiere mayor investigación sobre su biosíntesis y regulación (Block et al., 2019).
Avances en las últimas décadas, muestran que las plantas y los microorganismos han coevolucionado ejerciendo presiones selectivas mutuas y resultando en respuestas de defensa complejas (Pagán y García-Arenal, 2018; Upson et al., 2018). El área de estudio de las interacciones evolutivas moleculares (EvoMPMI, por sus siglas en inglés) busca integrar enfoques moleculares y evolutivos para descifrar estas complejas relaciones a lo largo del tiempo. Entender a fondo como las plantas ─a través de su MSP y exudados radicales ─ han evolucionado ante la presión de patógenos, brindaría herramientas adicionales para el diseño de estrategias de manejo integrado de plagas y enfermedades (Ponce de León y Montesano, 2017).
Límites del conocimiento y retos
A pesar de los avances, la mayoría de los fenómenos de defensa radical continúan siendo poco conocidos debido a la complejidad del subsuelo. El papel del ARN de interferencia en la interacción planta-patógeno (Rosa et al., 2018), la identificación de compuestos específicos que repelen o atraen organismos edáficos y la validación de extractos botánicos en ensayos de campo siguen siendo áreas que demandan estudio. Validar la eficacia de productos vegetales prometedores, como taninos, flavonoides y aceites esenciales ─bajo condiciones reales─ es un desafío clave, pues no siempre los resultados de laboratorio se replican en campo. En consecuencia, existe un amplio margen para la investigación interdisciplinaria que permita profundizar en estos mecanismos y aproveche el potencial de la rizósfera como un escenario principal de defensa y simbiosis (Biere y Goverse, 2016).
CONCLUSIONES
Las plantas enfrentan una amplia gama de patógenos y han desarrollado diversas estrategias de defensa basadas en metabolitos secundarios y otros mecanismos para limitar o evitar sus infecciones. No obstante, persisten interrogantes sobre el modo en que las raíces interpretan y emiten señales químicas frente a parásitos edáficos, incluidos los nematodos fitoparásitos, los microorganismos benéficos y otros factores del suelo El manejo sustentable demanda el uso inteligente de semioquímicos ya identificados y la exploración de nuevos compuestos que refuercen la defensa de las plantas y disminuyan la dependencia de plaguicidas químicos convencionales. Con una aproximación integral que abarque la biología molecular, la ecología química y la agronomía se podrán diseñar estrategias más eficaces para el manejo de plagas y enfermedades en cultivos. En este marco, los extractos botánicos que replican perfiles de exudados ofrecen una vía aplicada para traducir la ecología química de la rizósfera al manejo sustentable de Meloidogyne, sin perder el eje en los exudados radicales como punto de partida.
REFERENCIAS
Acevedo FE. 2020. Ecología química de interacciones entre plantas, insectos y controladores naturales de plagas herbívoras. In: Benavides P, Machado, Góngora CE, eds. El control natural de insectos en el ecosistema cafetero colombiano Cenicafé. https://doi.org/10.38141/10791/0001_5
Ayelo PM, Mohamed SA, Chailleux A, Yusuf AA, Pirk CWW, Deletre E. 2022. The parasitoid Dolichogenidea gelechiidivoris eavesdrops on semiochemicals from its host Tuta absoluta and tomato. Journal of Pest Science, 95(2), 633-652. https://doi.org/10.1007/s10340-021-01424-w
Bargali P, Kumar R, Devrani A, Mahawer SK, Karakoti H, Prakash O, Rawat S, Kumar S, Rawat DS, Dubey SK, Seth CS, Ahmad F. 2024. Essential oils work synergistically to mitigate pathogenic impact of Meloidogyne incognita, Rhizoctonia solani and Sclerotinia rolfsii. Biocatalysis and Agricultural Biotechnology, 60, 103160. https://doi.org/10.1016/j.bcab.2024.103160
Bavi K, Khavari-Nejad RA, Najafi F, Ghanati F. 2022. Phenolics and terpenoids change in response to yeast extract and chitosan elicitation in Zataria multiflora cell suspension culture. 3 Biotech, 12(8), 163. https://doi.org/10.1007/s13205-022-03235-x
Ben Mrid R, Benmrid B, Hafsa J, Boukcim H, Sobeh M, Yasri A. 2021. Secondary metabolites as biostimulant and bioprotectant agents: A review. Science of The Total Environment, 777, 146204. https://doi.org/10.1016/j.scitotenv.2021.146204
Biała-Leonhard, W., Zanin, L., Gottardi, S., de Brito Francisco, R., Venuti, S., Valentinuzzi, F., Mimmo, T, Cesco, S., Bassin, B., Martinoia, E., Pinton, R., Jasiński, M., Tomasi, N. (2021). Identification of an isoflavonoid transporter required for the nodule establishment of the rhizobium-fabaceae symbiotic interaction. Frontiers in Plant Science, 12. https://doi.org/10.3389/fpls.2021.758213
Biere A, Goverse A. 2016. Plant-mediated systemic interactions between pathogens, parasitic nematodes, and herbivores above- and belowground. Annual Review of Phytopathology, 54(1), 499-527. https://doi.org/10.1146/annurev-phyto-080615-100245
Blanco-Labra A, Aguirre-Mancilla C. 2002. Proteínas Involucradas en los Mecanismos de Defensa de Plantas. Acta Universitaria, (12)3, 3-28.
Blassioli-Moraes MC, Laumann RA, Michereff MFF, Borges M. 2019. Semiochemicals for integrated pest Management. In: Vaz Jr S, ed. Sustainable agrochemistry: A compendium of technologies. Cham: Springer International Publishing. https://doi.org/10.1007/978-3-030-17891-8_3.
Block AK, Vaughan MM, Schmelz EA, Christensen SA. 2019. Biosynthesis and function of terpenoid defense compounds in maize (Zea mays). Planta, 249(1), 21-30. https://doi.org/10.1007/s00425-018-2999-2
Boba A, Kostyn K, Kozak B, Wojtasik W, Preisner M, Prescha A, Gola EM, Lysh D, Dudek B, Szopa J, Kulma A. 2020. Fusarium oxysporum infection activates the plastidial branch of the terpenoid biosynthesis pathway in flax, leading to increased ABA synthesis. Planta, 251(2), 50. https://doi.org/10.1007/s00425-020-03339-9
Cantúa Ayala JA, Flores Olivas A, Valenzuela Soto JH. 2019. Compuestos orgánicos volátiles de plantas inducidos por insectos: situación actual en México. Revista mexicana de ciencias agrícolas, 10(3), 729-742. https://doi.org/10.29312/remexca.v10i3.678
Castro-Barquero L. 2024. Incremento de la fijación biológica de nitrógeno y el crecimiento de la soya (Glycine max) mediante el uso de hongos solubilizadores de fósforo. Revista Agronomía Costarricense, 48(1), 93-109. https://doi.org/10.15517/rac.v48i1.59136
Chamkhi I, El Omari N, Balahbib A, El Menyiy N, Benali T, Ghoulam C. 2022. Is the rhizosphere a source of applicable multi-beneficial microorganisms for plant enhancement? Saudi Journal of Biological Sciences, 29(2), 1246-1259. https://doi.org/10.1016/j.sjbs.2021.09.032
Chen N, Sun Q, Chen Z, Zhang X. 2025. Meloidogyne incognita significantly alters the cucumber root metabolome and enriches differential accumulated metabolites regulating nematode chemotaxis and infection. Horticulturae, 11(8), 892. https://doi.org/10.3390/horticulturae11080892
Chin S, Behm CA, Mathesius U. 2018. Functions of flavonoids in plant–nematode interactions. Plants, 7(4), 85. https://doi.org/10.3390/plants7040085
D’Addabbo T, Laquale S, Veronico P, Avato P, Argentieri MP. 2025. Nematicidal activity of the essential oil from Cinnamomum cassia and (E)-cinnamaldehyde against phytoparasitic nematodes. Journal of Pest Science, 98, 521–533. https://doi.org/10.1007/s10340-024-01816-8
Da Silva Sá GC, Bezerra PVV, da Silva MFA, da Silva LB, Barra PB, de Fátima Freire de Melo Ximenes M, Uchôa AF. 2023. Arbovirus vectors insects: are botanical insecticides an alternative for its management? Journal of Pest Science, 96(1), 1-20. https://doi.org/10.1007/s10340-022-01507-2
Dassanayake MK, Chong CH, Khoo T-J, Figiel A, Szumny A, Choo CM. 2021. Synergistic field crop pest management properties of plant-derived essential oils in combination with synthetic pesticides and bioactive molecules: A review. Foods, 10(9), 2016. https://doi.org/10.3390/foods10092016
Davison EK, Brimble MA. 2019. Natural product derived privileged scaffolds in drug discovery. Current Opinion in Chemical Biology, 52, 1-8. https://doi.org/10.1016/j.cbpa.2018.12.007
Degroote E, Schoorens C, Pockelé S, Stojilković B, Demeestere K, Mangelinckx S, Kyndt T. 2024. A combination of plant-based compounds and extracts acts nematicidal and induces resistance against Meloidogyne incognita in tomato. Frontiers in Plant Science, 15, 1411825. https://doi.org/10.3389/fpls.2024.1411825
De Carvalho GHF, de Andrade MA, de Araújo CN, Santos ML, de Castro NA, Charneau S, Monnerat R, de Santana J, Dourad Bastos IM. 2019. Larvicidal and pupicidal activities of eco-friendly phenolic lipid products from Anacardium occidentale nutshell against arbovirus vectors. Environmental Science and Pollution Research, 26(6), 5514-5523. https://doi.org/10.1007/s11356-018-3905-y
De-la-Peña C, Loyola-Vargas VM. 2014. Biotic interactions in the rhizosphere: A diverse cooperative enterprise for plant productivity. Plant Physiology, 166(2), 701–719. https://doi.org/10.1104/pp.114.241810
Dahlin P, Hallmann J. 2020. New insights on the role of allyl isothiocyanate in controlling the root knot nematode Meloidogyne hapla. Plants, 9(5), 603. https://doi.org/10.3390/plants9050603
Dou X, Wu Y, Luo J, Yin X, Fu M, Zeng X, Wang H. 2024. Activity of the ethanolic extract obtained from Citrus microcarpa pericarps against Meloidogyne enterolobii, and chemical composition analysis. HortScience, 59(6), 858–864. https://doi.org/10.21273/HORTSCI17817-24
Elsharkawy MM, Al-Askar AA, Behiry SI, Abdelkhalek A, Saleem MH, Kamran M, Derbalah A. 2022. Resistance induction and nematicidal activity of certain monoterpenes against tomato root-knot caused by Meloidogyne incognita. Frontiers in Plant Science, 13, 982414. https://doi.org/10.3389/fpls.2022.982414
El-Saadony MT, Abuljadayel DA, Shafi ME, Albaqami NM, Desoky E-SM, El-Tahan AM, Mesiha PK, Elnahal ASM, Almakas A, Taha AE, El-Mageed TAA, Hassanin AA, Elrys AS, Saad AM 2021. Control of foliar phytoparasitic nematodes through sustainable natural materials: Current progress and challenges. Saudi Journal of Biological Sciences, 28(12), 7314-7326. https://doi.org/10.1016/j.sjbs.2021.08.035
Eugui D, Escobar C, Velasco P, Poveda J. 2022. Glucosinolates as an effective tool in plant-parasitic nematodes control: Exploiting natural plant defenses. Applied Soil Ecology, 176, 104497. https://doi.org/10.1016/j.apsoil.2022.104497
Fanelli E, Vovlas A, D’Addabbo T, De Luca F. 2025. Molecular mechanism of Cinnamomum zeylanicum and Citrus aurantium essential oils against the root-knot nematode, Meloidogyne incognita. Scientific Reports, 15, 6077. https://doi.org/10.1038/s41598-025-90529-8
Campo Fernández M, Ambuludí Fárez DL, Cepeda Roblez NC, Márquez Hernández I, San Martín Galván D, Cuesta Rubio O. 2018. Composición química y actividad antimicrobiana del extracto etanólico de las partes aéreas de Minthostachys mollis Griseb. Revista Cubana de Farmacia, 51(2). https://revfarmacia.sld.cu/index.php/far/article/view/221
Gahukar RT. 2012. Evaluation of plant-derived products against pests and diseases of medicinal plants: A review. Crop Protection, 42, 202-209. https://doi.org/10.1016/j.cropro.2012.07.026
Gautier C, Montarry J, Piriou C, Renault L, Porte C, Yvin J-C, Nguema-Ona E, Fournet S. 2021. Impact of the genetic diversity of three cyst nematodes on the effectiveness of root exudates to induce hatching. European Journal of Plant Pathology, 161(3), 553-563. https://doi.org/10.1007/s10658-021-02341-5
Guerrieri A, Floková K, Vlaar LE, Schilder ML, Kramer G, Chojnacka A, van Dijk YR, Bouwmeester HJ, Dong L. 2021. UPLC-MS/MS analysis and biological activity of the potato cyst nematode hatching stimulant, solanoeclepin A, in the root exudate of Solanum spp. Planta, 254(6), 112. https://doi.org/10.1007/s00425-021-03766-2
Hakim S, Naqqash T, Nawaz MS, Laraib I, Siddique MJ, Zia R, Mirza MS, Imran A. 2021. Rhizosphere engineering with plant growth-promoting microorganisms for agriculture and ecological sustainability. Frontiers in Sustainable Food Systems, 5. https://doi.org/10.3389/fsufs.2021.617157
Haroun HM, Aydınlı G, Mennan S. 2022. Screening of the nematicidal potential of some spice extracts against root-knot nematode, Meloidogyne arenaria (Neal, 1889) Chitwood, 1949 (Tylenchida: Meloidogynidae). Turkish Journal of Entomology, 46(3), 275–288. https://doi.org/10.16970/entoted.1107144
Hawes M, Allen C, Turgeon BG, Curlango-Rivera G, Tran TM, Huskey DA, Xiong Z. 2016. Root border cells and their role in plant defense. Annual Review of Phytopathology, 54(1), 143-161. https://doi.org/10.1146/annurev-phyto-080615-100140
Hazafa A, Rehman K-U, Jahan N, Jabeen Z. 2020. The role of polyphenol (flavonoids) compounds in the treatment of cancer cells. Nutrition and Cancer, 72(3), 386-397. https://doi.org/10.1080/01635581.2019.1637006
He Y, Xu J, Wang X, He X, Wang Y, Zhou J, Zhang S, Meng X. 2019. The Arabidopsis Pleiotropic Drug Resistance Transporters PEN3 and PDR12 mediate camalexin secretion for resistance to Botrytis cinerea. The Plant Cell, 31(9), 2206-2222. https://doi.org/10.1105/tpc.19.00239
Holbein J, Franke RB, Marhavý P, Fujita S, Górecka M, Sobczak M, Geldne N, Schreiber L, Grundler FMW, Siddique S. 2019. Root endodermal barrier system contributes to defence against plant-parasitic cyst and root-knot nematodes. The Plant Journal, 100(2), 221-236. https://doi.org/10.1111/tpj.14459
Holopainen JK, Virjamo V, Ghimire RP, Blande JD, Julkunen-Tiitto R, Kivimäenpää M. 2018. Climate change effects on secondary compounds of forest trees in the northern hemisphere. Frontiers in Plant Science, 9. https://doi.org/10.3389/fpls.2018.01445
Hu L, Robert CAM, Cadot S, Zhang X, Ye M, Li B, Manzo D, Chervet N, Steinger T, van der Heijden MGA, Schlaeppi K, Erb M. 2018. Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nature Communications, 9(1), 2738. https://doi.org/10.1038/s41467-018-05122-7
Jamshidi-Kia F, Lorigooini Z, Amini-Khoei H. 2018. Medicinal plants: Past history and future perspective. Journal of herbmed pharmacology, 7(1). http://doi.org/10.15171/jhp.2018.01
Jan R, Asaf S, Numan M, Lubna, Kim K-M. 2021. Plant secondary metabolite miosynthesis and transcriptional regulation in response to biotic and abiotic stress sonditions. Agronomy, 11(5), 968. https://doi.org/10.3390/agronomy11050968
Jardim IN, Oliveira DF, Campos VP, Freire ES, Silva GH, Nunes AS. 2020. Garlic essential oil reduces the population of Meloidogyne incognita in tomato plants. European Journal of Plant Pathology, 157(2), 197–209. https://doi.org/10.1007/s10658-020-02000-1
Kapoor B, Gulati M, Gupta R, Singh SK, Gupta M, Nabi A, Chawla PA. 2021. A review on plant flavonoids as potential anticancer agents. Current Organic Chemistry, 25(6), 737-747. https://doi.org/10.2174/1385272824999201126214150
Kessler A, Kalske A. 2018. Plant secondary metabolite diversity and species interactions. Annual Review of Ecology, Evolution, and Systematics, 49(1), 115-138. https://doi.org/10.1146/annurev-ecolsys-110617-062406
Khalid M, Saeed ur R, Bilal M, Huang D-F. 2019. Role of flavonoids in plant interactions with the environment and against human pathogens — A review. Journal of Integrative Agriculture, 18(1), 211-230. https://doi.org/10.1016/S2095-3119(19)62555-4
Kirwa HK, Murungi LK, Beck JJ, Torto B. 2018. Elicitation of differential responses in the root-knot nematode Meloidogyne incognita to tomato root exudate cytokinin, flavonoids, and alkaloids. Journal of Agricultural and Food Chemistry, 66(43), 11291-11300. https://doi.org/10.1021/acs.jafc.8b05101
Korenblum E, Massalha H, Aharoni A. 2022. Plant–microbe interactions in the rhizosphere via a circular metabolic economy. The Plant Cell Retrieved from https://doi.org/10.1093/plcell/koac163
Lengai GMW, Muthomi JW, Mbega ER. 2020. Phytochemical activity and role of botanical pesticides in pest management for sustainable agricultural crop production. Scientific African, 7, e00239. https://doi.org/10.1016/j.sciaf.2019.e00239
Liu W, Park S-W. 2018. Underground mystery: Interactions between plant roots and parasitic nematodes. Current Plant Biology, 15, 25-29. https://doi.org/10.1016/j.cpb.2018.11.004
López-Cardona N, López-Casallas M. (2018). Evaluación de resistencia de cultivares de soya (Glycine max L.) a aislamientos de Phytophthora sojae Kaufm. & Gerd. de la altillanura colombiana. Revista De La Academia Colombiana De Ciencias Exactas, Físicas Y Naturales, 42(164), 262-268. https://doi.org/10.18257/raccefyn.695
Lu X, Zhang L, Huang W, Zhang S, Zhang S, Li F, Zhang H, Sun R, Zhao J, Li G. 2022. Integrated volatile metabolomics and transcriptomics analyses reveal the influence of infection TuMV to volatile organic compounds in Brassica rapa. Horticulturae, 8(1), 57. https://doi.org/10.3390/horticulturae8010057
Mezerket A, Palomares-Rius JE, Bouasla S, Saib H. 2025. Phytochemical profile and nematicidal potential of essential oil from Algerian wild Origanum vulgare subsp. glandulosum Desf. Turkish Journal of Biology, 49(1), 52–59. https://doi.org/10.55730/1300-0152.2723
Mnyambo NM, Rantho LP, Dube ZP, Timana M. 2024. Timing of plant extracts application in the management of Meloidogyne incognita on tomato plants. International Journal of Plant Biology, 15(4), 1108–1117. https://doi.org/10.3390/ijpb15040077
Ngala B, Dewaegeneire P, Robilliard E, Mariette N, Manceau F, Denis M-C, Porte C, Neveux M-S, Le Roux A-C, Fournet S, Montarry J. 2024. Lure and starve: host root exudates to suppress field populations of cyst nematodes. Applied Soil Ecology, 201, 105490. https://doi.org/10.1016/j.apsoil.2024.105490
Ntalli N, Zochios G, Nikolaou P, Winkiel M, Petrelli R, Bonacucina G, Perinelli DR, Spinozzi E, Maggi F, Benelli G. 2023. Carlina acaulis essential oil nanoemulsion for managing Meloidogyne incognita. Industrial Crops and Products, 193, 116180. https://doi.org/10.1016/j.indcrop.2022.116180
Martínez-Bastidas TF, Romero-Castillo RA, Amarillas-Bueno LA, López-Meyer M, Ramírez K, Sañudo-Barajas JA, Osuna-Enciso T, Heredia JB, Lightbourn-Rojas LA, León-Félix J. 2017. Proteínas G heterotriméricas: Señalización de plantas en condiciones de estrés ambiental. Revista Fitotecnia Mexicana, 40(2), 169-180. https://doi.org/10.35196/rfm.2017.2.169-180
Más Toro D, Martínez Aguilar Y, Rodríguez Bertot R, Pupo Torres G, Rosabal Nava O, Olmo González C. 2017. Análisis preliminar de los metabolitos secundarios de polvos mixtos de hojas de plantas medicinales. Revista Cubana de Plantas Medicinales, 22, 0-0. http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S1028-47962017000100005&nrm=iso
Mathesius U, Costa SR. 2021. Plant signals differentially affect rhizosphere nematode populations. Journal of Experimental Botany, 72(10), 3496-3499. https://doi.org/10.1093/jxb/erab149
Meents AK, Mithöfer A. 2020. Plant–plant communication: Is there a role for volatile damage-associated molecular patterns? Frontiers in Plant Science, 11. https://doi.org/10.3389/fpls.2020.583275
Menéndez E, Navarro J, López J, Dalmau A. 2020. Mecanismos de defensa en plantas. Proteínas relacionadas con la patogenicidad. Revista de Investigaciones de la Universidad Le Cordon Blue, 7(2), 98-109. https://doi.org/10.36955/RIULCB.2020v7n2.010
Menikheim CB, Mousavi S, Bereswill S, Heimesaat MM. 2024. Polyphenolic compounds in the combat of foodborne infections – An update on recent evidence. European Journal of Microbiology & Immunology, 14(2), 116-125. https://doi.org/10.1556/1886.2024.00018
Miller SA. Ferreira JP, LeJeune JT. 2022. Antimicrobial Use and Resistance in Plant Agriculture: A One Health Perspective. Agriculture, 12, 289. https://doi.org/ 10.3390/agriculture12020289
Monzote L, García J, González R, Scotti MT, Setzer WN. 2021. Bioactive Essential Oils from Cuban Plants: An Inspiration to Drug Development. Plants, 10(11), 2515. https://doi.org/10.3390/plants10112515
Nguyen NH, Trotel-Aziz P, Clément C, Jeandet P, Baillieul F, Aziz A. 2022. Camalexin accumulation as a component of plant immunity during interactions with pathogens and beneficial microbes. Planta, 255(6), 116. https://doi.org/10.1007/s00425-022-03907-1
Ochola J, Coyne D, Cortada L, Haukeland S, Ng'ang'a M, Hassanali A, Opperman C, Torto B. 2021. Cyst nematode bio-communication with plants: implications for novel management approaches. Pest Management Science, 77(3), 1150-1159. https://doi.org/10.1002/ps.6105
Oosterbeek M, Lozano-Torres JL, Bakker J, Goverse A. 2021. Sedentary plant-parasitic nematodes alter auxin homeostasis via multiple strategies. Frontiers in Plant Science, 12. https://doi.org/10.3389/fpls.2021.668548
Pagán I., García-Arenal F. 2018. Tolerance to plant pathogens: Theory and experimental evidence. International Journal of Molecular Sciences, 19(3), 810. https://doi.org/10.3390/ijms19030810
Palomares-Rius JE, Hasegawa K, Siddique S, Vicente CSL. 2021. Editorial: Protecting our crops - approaches for plant parasitic nematode control. Frontiers in Plant Science, 12. https://doi.org/10.3389/fpls.2021.726057
Park Y-S, Ryu C-M. 2021. Understanding Plant Social Networking System: Avoiding Deleterious Microbiota but Calling Beneficials. International Journal of Molecular Sciences, 22(7), 3319. https://doi.org/10.3390/ijms22073319
Perczak A, Gwiazdowska D, Marchwińska K, Juś K, Gwiazdowski R, Waśkiewicz A. 2019. Antifungal activity of selected essential oils against Fusarium culmorum and F. graminearum and their secondary metabolites in wheat seeds. Archives of Microbiology, 201(8), 1085-1097. https://doi.org/10.1007/s00203-019-01673-5
Ponce de León I, Montesano M. 2017. Adaptation mechanisms in the evolution of moss defenses to microbes. Frontiers in Plant Science, 8. https://doi.org/10.3389/fpls.2017.00366
Ramya M, Jang S, An H-R, Lee S-Y, Park P-M, Park PH. 2020. Volatile organic compounds from orchids: From synthesis and function to gene regulation. International Journal of Molecular Sciences, 21(3), 1160. https://doi.org/10.3390/ijms21031160
Rolfe SA, Griffiths J, Ton J. 2019. Crying out for help with root exudates: adaptive mechanisms by which stressed plants assemble health-promoting soil microbiomes. Current Opinion in Microbiology, 49, 73-82. https://doi.org/10.1016/j.mib.2019.10.003
Rosa C, Kuo Y-W, Wuriyanghan H, Falk BW. 2018. RNA interference mechanisms and applications in plant pathology. Annual Review of Phytopathology, 56(1), 581-610. https://doi.org/10.1146/annurev-phyto-080417-050044
Roy A, Khan A, Ahmad I, Alghamdi S, Rajab BS, Babalghith AO, Alshahrani MY, Islam S, Islam MR. 2022. Flavonoids a bioactive compound from medicinal plants and its therapeutic applications. BioMed Research International, 2022, 5445291. https://doi.org/10.1155/2022/5445291
Schwerdtner U, Spohn M. 2022. Plant species interactions in the rhizosphere increase maize N and P acquisition and maize yields in intercropping. Journal of Soil Science and Plant Nutrition, 22, 3868-3884. https://doi.org/10.1007/s42729-022-00936-3
Seiml-Buchinger VV, Zinovieva SV, Udalova ZV, Matveeva EM. 2019. Jasmonic acid modulates Meloidogyne incognita – tomato plant interactions. Nematology, 21(2), 171-180. https://doi.org/10.1163/15685411-00003205
Shai KN, Chakale MV, Matarechera SA, Amoo SO. 2024. Utilisation of botanicals for the management of pests and diseases affecting crops in sub-Saharan Africa: A review. Journal of Natural Pesticide Research, 7, 100066. https://doi.org/10.1016/j.napere.2023.100066
Shen N, Wang T, Gan Q, Liu S, Wang L, Jin B. 2022. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chemistry, 383, 132531. https://doi.org/10.1016/j.foodchem.2022.132531
Sikder MM, Vestergård M, Kyndt T, Kudjordjie EN, Nicolaisen M. 2021. Phytohormones selectively affect plant parasitic nematodes associated with Arabidopsis roots. New Phytologist, 232(3), 1272-1285. https://doi.org/10.1111/nph.17549
Sousa IP, Chagas-Paula DA, Tiossi RFJ, Silva EdO, Miranda MA, de Oliveira RB, Miranda AM, Barbosa de Oliveira R, Cropanese Spadaro AC, Bastos JK, Cardoso Furtado NAJ, Da Costa FB. 2019. Essential oils from Tithonia diversifolia display potent anti-oedematogenic effects and inhibit acid production by cariogenic bacteria. Journal of Essential Oil Research, 31(1), 43-52. https://doi.org/10.1080/10412905.2018.1500315
Spiegler V, Liebau E, Hensel A. 2017. Medicinal plant extracts and plant-derived polyphenols with anthelmintic activity against intestinal nematodes. Natural Product Reports, 34(6), 627-643. http://dx.doi.org/10.1039/C6NP00126B
Thakur M, Bhattacharya S, Khosla PK, Puri S. 2019. Improving production of plant secondary metabolites through biotic and abiotic elicitation. Journal of Applied Research on Medicinal and Aromatic Plants, 12, 1-12. https://doi.org/10.1016/j.jarmap.2018.11.004
Tlak Gajger I, Dar SA. 2021. Plant allelochemicals as sources of insecticides. Insects, 12(3), 189. https://doi.org/10.3390/insects12030189
Torto B, Cortada L, Murungi LK, Haukeland S, Coyne DL. 2018. Management of cyst and root knot nematodes: A chemical ecology perspective. Journal of Agricultural and Food Chemistry, 66(33), 8672-8678. https://doi.org/10.1021/acs.jafc.8b01940
Upson JL, Zess EK, Białas A, Wu C-h, Kamoun S. 2018. The coming of age of EvoMPMI: evolutionary molecular plant–microbe interactions across multiple timescales. Current Opinion in Plant Biology, 44, 108-116. https://doi.org/10.1016/j.pbi.2018.03.003
Vélez-Ruiz MC, Meza-Vera RJ, Abasolo-Pacheco F, Álvarez-Romero PI. (2022). Use of botanical extracts to control the aphid (Myzus persicae: Aphididae) and whitef ly (Bemisia tabaci: Aleyrodidae) in pepper crop (Capsicum anuum: Solanaceae), in Ecuador. REVISTA TERRA LATINOAMERICANA, 40. https://doi.org/10.28940/terra.v40i0.1454
Verrillo M, Cozzolino V, Spaccini R, Piccolo A. 2021. Humic substances from green compost increase bioactivity and antibacterial properties of essential oils in Basil leaves. Chemical and Biological Technologies in Agriculture, 8(1), 28. https://doi.org/10.1186/s40538-021-00226-7
Vimala G, Machal M, Rana VS, Gowda APA, Kumar V, Shakil NA, Pervez R, Singh AK, Kumar R, Jaiman M, Pankaj. 2025. Effect of botanicals, organic nutrient sources, and bio-control agents on root-knot nematode (Meloidogyne incognita) infecting tomato. Frontiers in Plant Science, 16, 1602326. https://doi.org/10.3389/fpls.2025.1602326
Wen W, Alseekh S, Fernie AR. 2020. Conservation and diversification of flavonoid metabolism in the plant kingdom. Current Opinion in Plant Biology, 55, 100-108. https://doi.org/10.1016/j.pbi.2020.04.004
Wink M. 2015. Modes of action of herbal medicines and plant secondary metabolites. Medicines, 2(3), 251-286. https://doi.org/10.3390/medicines2030251
Yadav B, Jogawat A, Rahman MS, Narayan OP. 2021. Secondary metabolites in the drought stress tolerance of crop plants: A review. Gene Reports, 23, 101040. https://doi.org/10.1016/j.genrep.2021.101040
Zélé F, Magalhães S, Kéfi S, Duncan AB. 2018. Ecology and evolution of facilitation among symbionts. Nature Communications, 9(1), 4869. https://doi.org/10.1038/s41467-018-06779-w
Zhang H, Yang Y, Mei X, Li Y, Wu J, Li Y, Wang H, Huang H, Yang M, He X, Zhu S, Liu Y. 2020. Phenolic acids released in maize rhizosphere during maize-soybean intercropping inhibit Phytophthora Blight of soybean. Frontiers in Plant Science, 11. https://doi.org/10.3389/fpls.2020.00886